

Crosstalk between cancer-associated fibroblasts and non-neuroendocrine tumor cells in small cell lung cancer involves in glycolysis and antigen-presenting features
- PMID: 39722014
- PMCID: PMC11669202
- DOI: 10.1186/s10020-024-01051-y
Crosstalk between cancer-associated fibroblasts and non-neuroendocrine tumor cells in small cell lung cancer involves in glycolysis and antigen-presenting features
Abstract
Background: Small cell lung cancer (SCLC) is a highly fatal malignancy, the complex tumor microenvironment (TME) is a critical factor affecting SCLC progression. Cancer-associated fibroblasts (CAFs) are crucial components of TME, yet their role in SCLC and the underlying mechanisms during their interaction with SCLC cells remain to be determined.
Methods: Microenvironmental cell components were estimated using transcriptome data from SCLC tissue available in public databases, analyzed with bioinformatic algorithms. A co-culture system comprising MRC5 fibroblasts and SCLC cell lines was constructed. RNA sequencing (RNA-seq) was performed on co-cultured and separately cultured MRC5 and H196 cells to identify differentially expressed genes (DEGs) and enriched signaling pathways. Glycolysis and STING signaling in SCLC cells were assessed using glucose uptake assays, qRT-PCR, and Western blot analysis. Immunohistochemical staining of SCLC tissue arrays quantified α-SMA, HLA-DRA and CD8 expression.
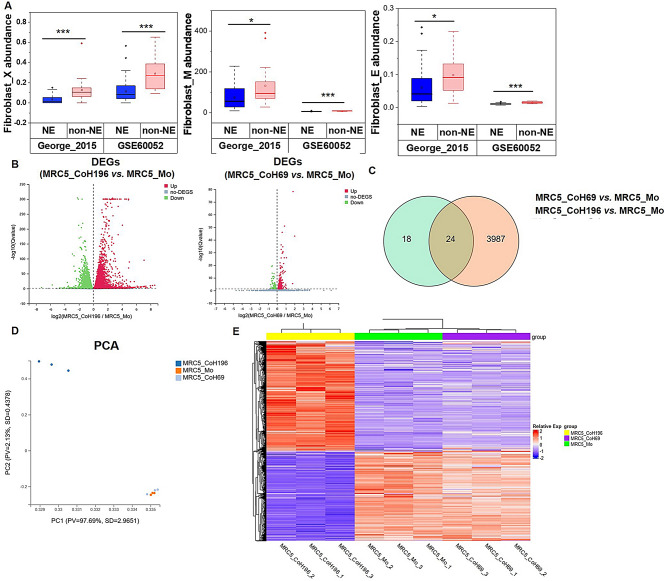
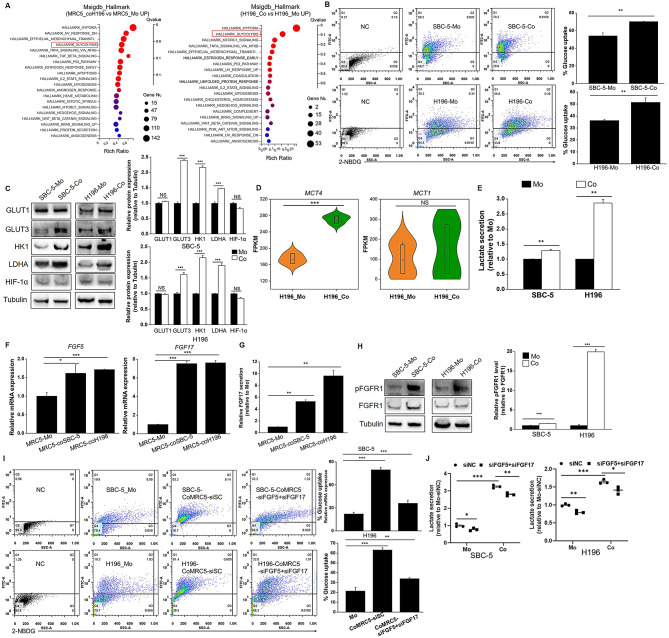
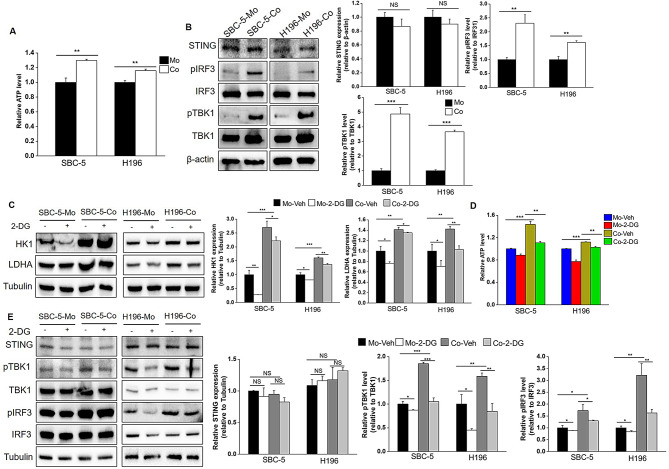
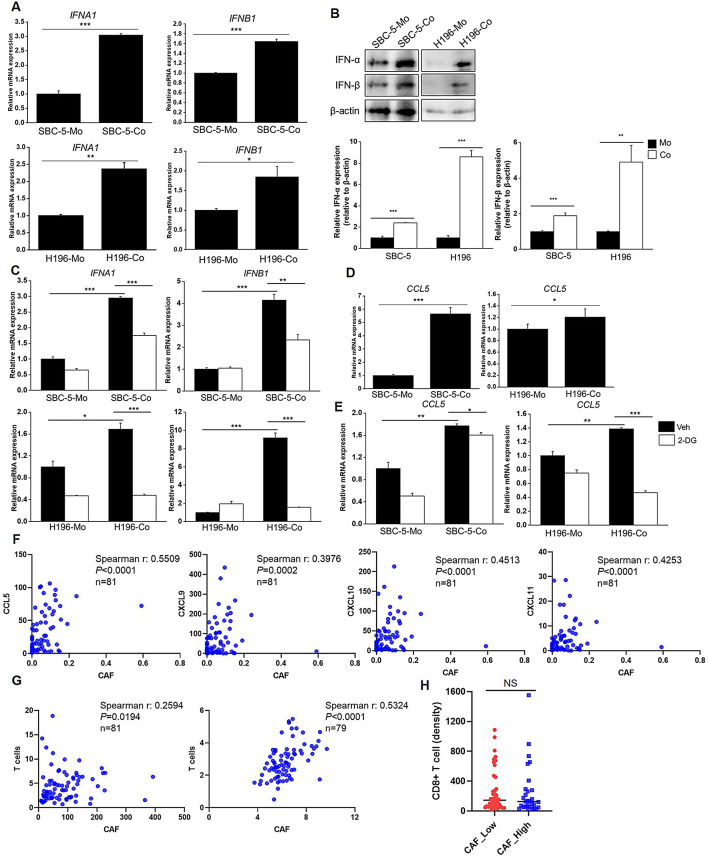
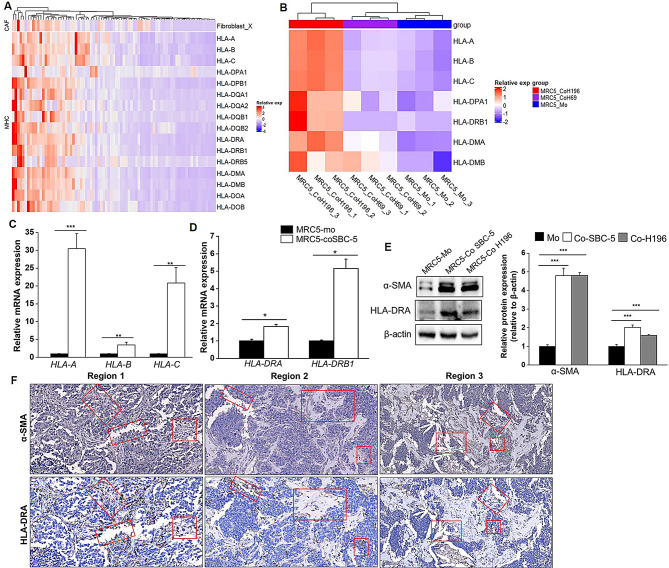
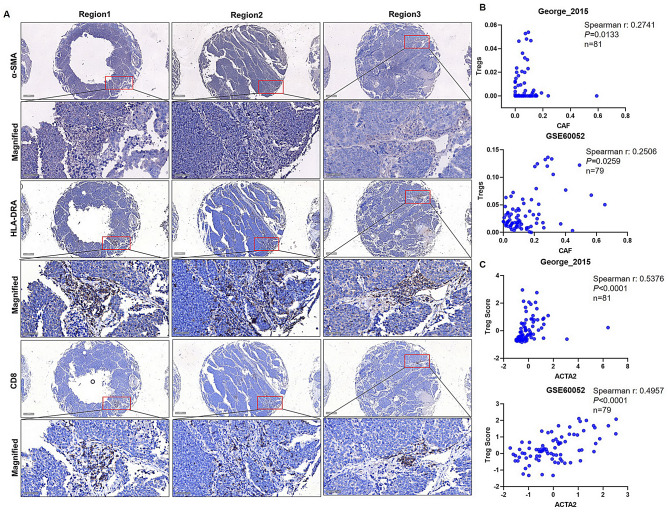
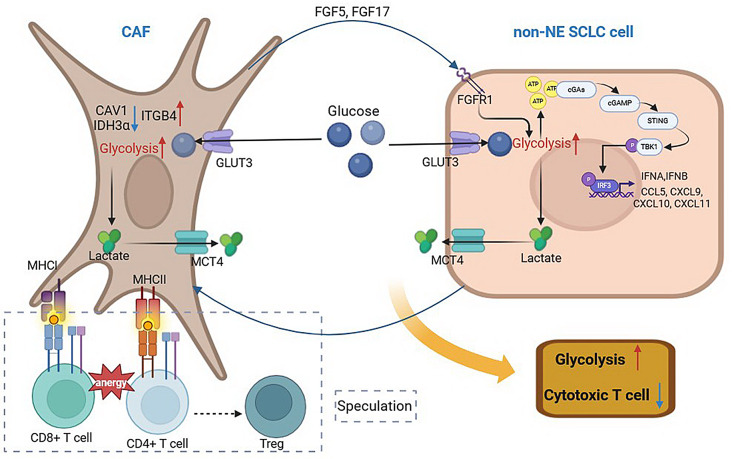
Results: Non-neuroendocrine (non-NE) SCLC-derived CAFs exhibited more abundance and DEGs than NE SCLC-derived CAFs did, which interact with non-NE SCLC cells can induce the enrichment of glycolysis-related genes, increasement of glucose uptake, upregulation of glycolytic signaling proteins in non-NE SCLC cells and accumulation of lactate in the extracellular environment, confirming CAF-mediated glycolysis promotion. Additionally, glycolysis-induced ATP production activated STING signaling in non-NE SCLC cells, which upregulated T cell chemo-attractants. However, CAF abundance did not correlate with CD8 + T cell numbers in SCLC tissues. Additionally, non-NE SCLC cell-educated CAFs exhibited features of antigen-presenting CAFs (apCAFs), as indicated by the expression of major histocompatibility complex (MHC) molecules. Co-localization of HLA-DRA and α-SMA signals in SCLC tissues confirmed apCAF presence. The apCAFs and CD8 + T cells were co-located in the SCLC stroma, and there was a positive correlation between CAFs and regulatory T cell (Treg) abundance.
Conclusion: Our findings suggest that crosstalk between CAFs and non-NE SCLC cells promotes glycolysis in non-NE SCLC cells, thereby increase T cell chemo-attractant expression via activating STING signaling. On the other hand, it promotes the presence of apCAFs, which probably contributes to CD8 + T cell trapping and Treg differentiation. This study emphasizes the pro-tumor function of CAFs in SCLC by promoting glycolysis and impairing T cell function, providing direction for the development of novel therapeutic approaches targeting CAF in SCLC.
Keywords: Antigen presentation; CAF; Glycolysis; SCLC; STING signaling.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: The tissue arrays were obtained from ZK bioaitech company, informed patient consent was obtained prior to collection. The study was approved by Ethics Committee of Changsha Yaxiang Biotechnology Co., Ltd (Csyayj20231120). All methods were performed as per the relevant regulations and the Declaration of Helsinki. Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures
References
MeSH terms
Grants and funding
- 2022JC008/Health Commission of Jilin Province
- 2023 Post-doctoral funding program/Department of Human Resources and Social Security of Jilin Provincial
- 20240304040SF/Department of Science and Technology of Jilin Province
- YDZ202401114ZYTS/Department of Science and Technology of Jilin Province
- YDZJ202301ZYTS512/Department of Science and Technology of Jilin Province
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
