

Transcriptome Analysis Reveals Norathyriol Prolongs the Lifespan via Regulating Metabolism in C. elegans
- PMID: 39728497
- PMCID: PMC11678140
- DOI: 10.3390/metabo14120716
Transcriptome Analysis Reveals Norathyriol Prolongs the Lifespan via Regulating Metabolism in C. elegans
Abstract
Background: Aging and age-related diseases are closely linked to an imbalance in energy supply and demand, a condition that can potentially be mitigated through various interventions, including the use of naturally occurring molecules. Norathyriol (NL), a tetrahydroxyxanthone compound, is prevalent in mango fruit and medicinal plants. While studies have indicated that NL may influence metabolism, its effects on aging have not been extensively explored.
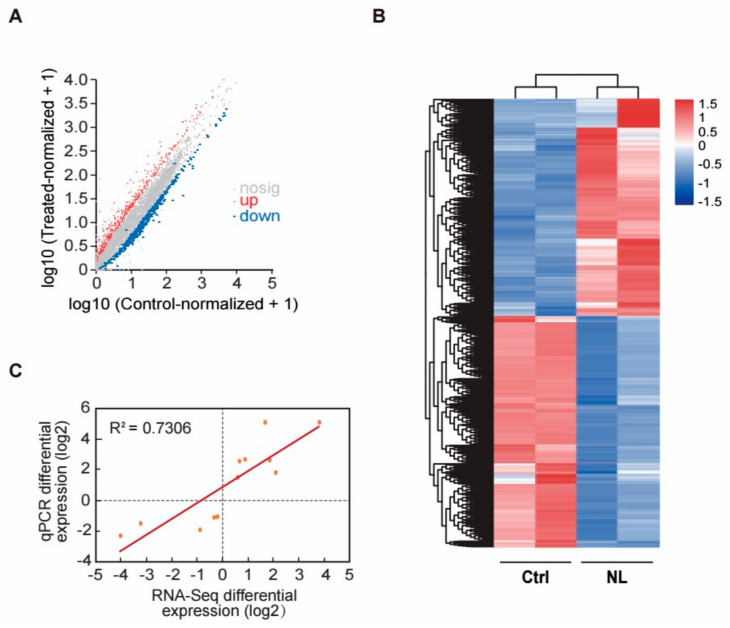
Methods: We conducted lifespan analysis and measured lipofuscin accumulation in C. elegans model to evaluate the effects of NL on aging. Additionally, we identified differentially expressed genes (DEGs) through comprehensive RNA-sequencing (RNA-seq) analysis and performed gene ontology (GO) and kyoto encyclopedia of genes and genomes (KEGGs) pathway analyses to elucidate the molecular mechanisms underlying NL's effects.
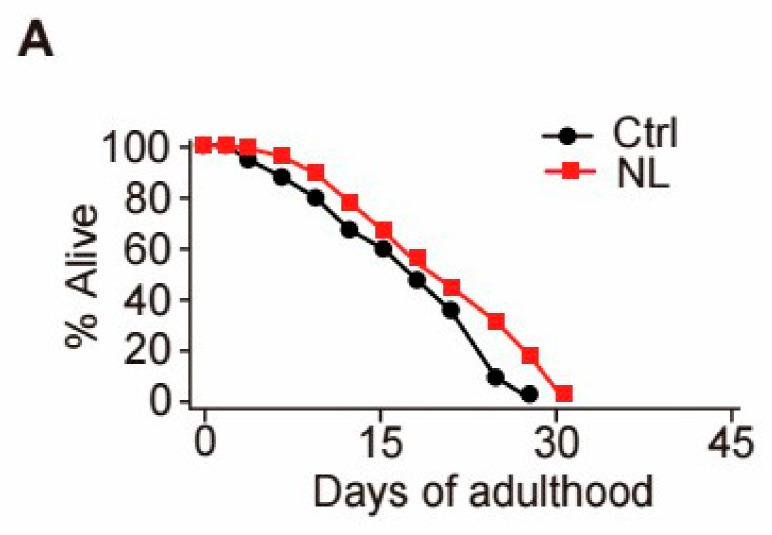
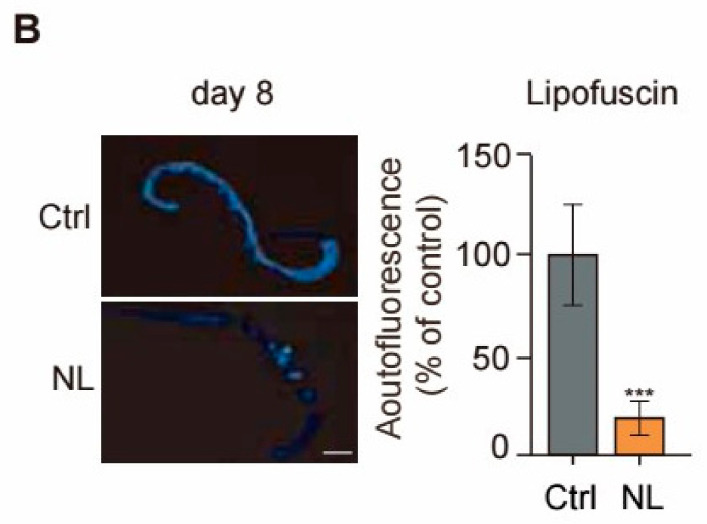
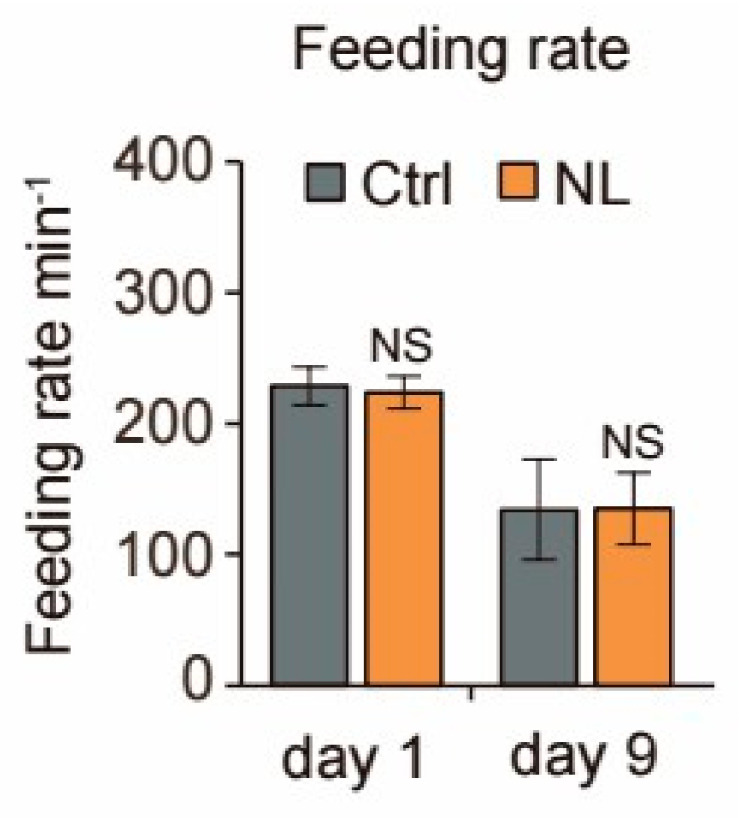
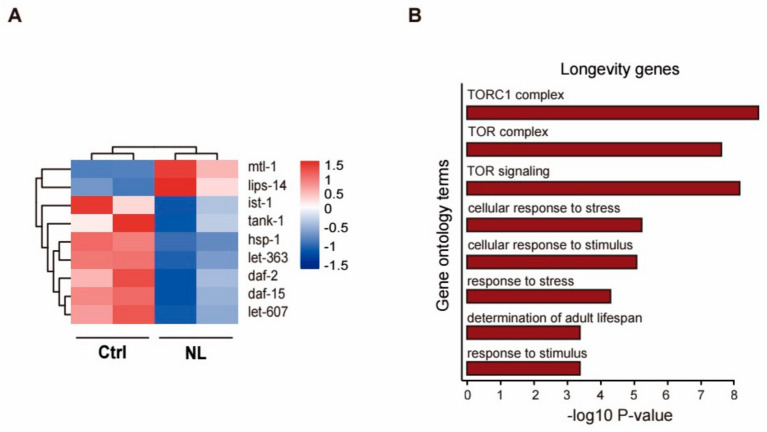
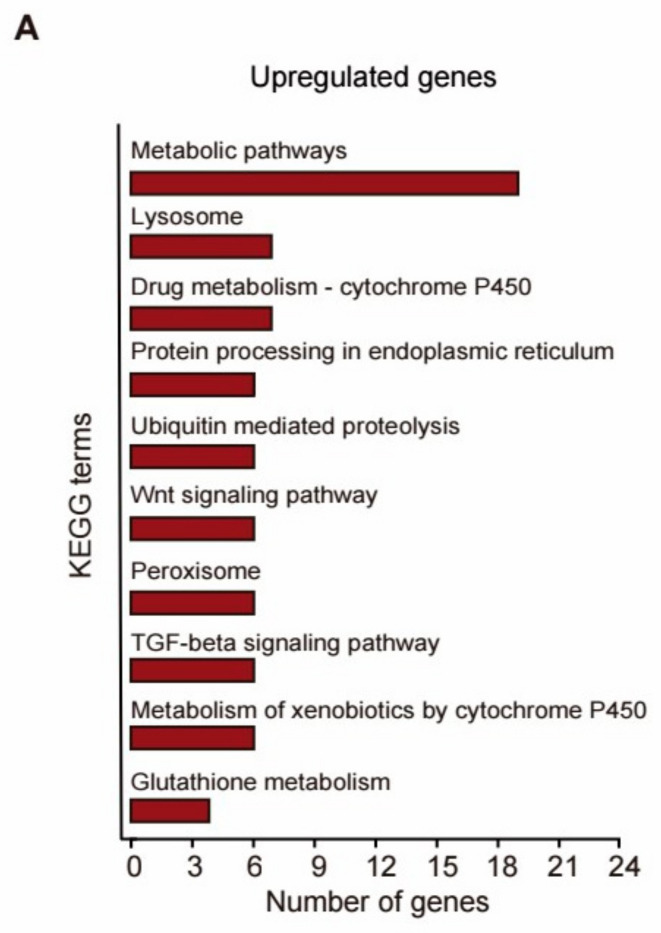
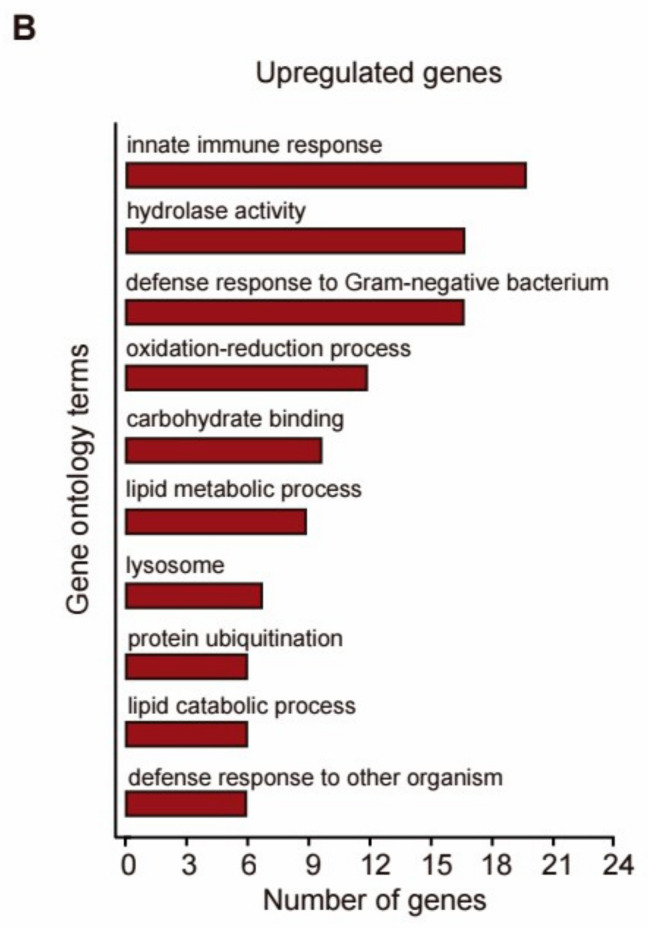
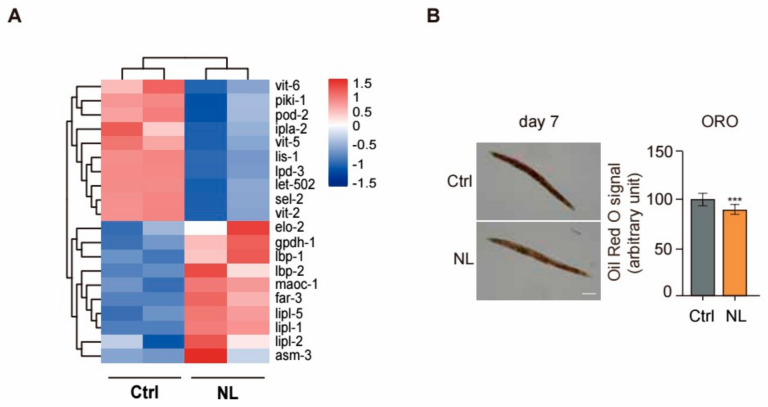
Results: Our study demonstrated that NL at 50 μM extends the lifespan by 15.9% and reduces lipofuscin accumulation in C. elegans without impacting their feeding capabilities. A total of 928 DEGs were identified in NL-treated worms. The analysis of DEGs indicated that NL's longevity-promoting effects might be due to its regulation of gene expression in lipid metabolism and immune response pathways. Furthermore, the insulin/insulin-like growth factor (IGF)-1 and target of rapamycin (TOR) signaling pathways were implicated in the lifespan-extending effect of NL.
Conclusions: These findings broaden the bioactivity profile of polyphenols and highlight the need for further investigation into the therapeutic potential of NL in combating age-related diseases.
Keywords: C. elegans; aging; lipid metabolism; norathyriol; transcriptome analysis.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Astaxanthin Induces Transcriptomic Responses Associated with Lifespan Extension in Caenorhabditis elegans.Antioxidants (Basel). 2022 Oct 27;11(11):2115. doi: 10.3390/antiox11112115. Antioxidants (Basel). 2022. PMID: 36358487 Free PMC article.
-
Ether lipid biosynthesis promotes lifespan extension and enables diverse pro-longevity paradigms in Caenorhabditis elegans.Elife. 2023 Aug 22;12:e82210. doi: 10.7554/eLife.82210. Elife. 2023. PMID: 37606250 Free PMC article.
-
Phloretic acid requires the insulin/IGF-1 pathway and autophagy to enhance stress resistance and extend the lifespan of Caenorhabditis elegans.Front Pharmacol. 2024 Mar 27;15:1384227. doi: 10.3389/fphar.2024.1384227. eCollection 2024. Front Pharmacol. 2024. PMID: 38601465 Free PMC article.
-
Control of aging by the renin-angiotensin system: a review of C. elegans, Drosophila, and mammals.Front Pharmacol. 2022 Sep 14;13:938650. doi: 10.3389/fphar.2022.938650. eCollection 2022. Front Pharmacol. 2022. PMID: 36188619 Free PMC article. Review.
-
Key proteins and pathways that regulate lifespan.J Biol Chem. 2017 Apr 21;292(16):6452-6460. doi: 10.1074/jbc.R116.771915. Epub 2017 Mar 6. J Biol Chem. 2017. PMID: 28264931 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Miscellaneous
