

Image Analysis and Untargeted Metabolomics Reveal Potential Phytotoxins from Fusarium venenatum Against Major Parasitic Weed Phelipanche ramosa (L.) Pomel
- PMID: 39728789
- PMCID: PMC11678980
- DOI: 10.3390/toxins16120531
Image Analysis and Untargeted Metabolomics Reveal Potential Phytotoxins from Fusarium venenatum Against Major Parasitic Weed Phelipanche ramosa (L.) Pomel
Abstract
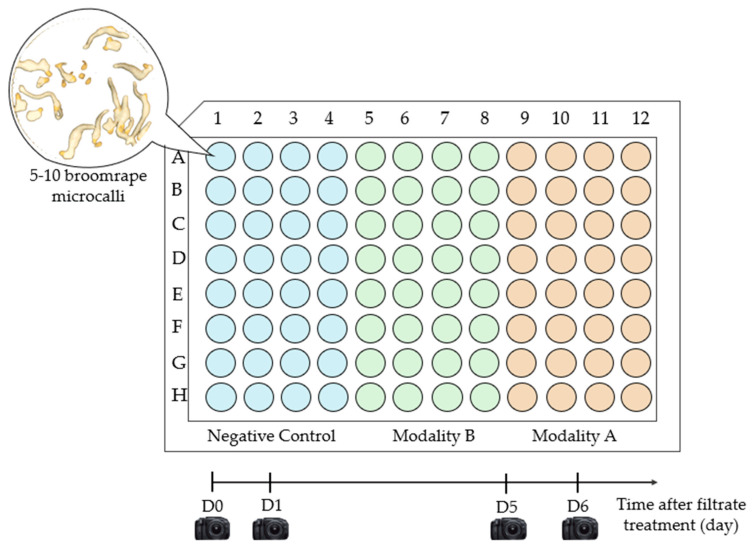
Branched broomrape (Phelipanche ramosa (L.) Pomel), an obligate parasitic weed with a wide host range, is known for its devasting effects on many crops worldwide. Soil fungi, notably Fusarium sp., are described as pathogenic to broomrape, while the hypothesis of the phytotoxicity of fusaric acid produced by F. verticillioides for parasitic weeds of the genus Orobanche has been proposed. Using image analysis and untargeted metabolomics, this study investigated fungal metabolites phytotoxic for P. ramosa and produced by the F. venenatum MIAE02836 strain, isolated from symptomatic broomrapes and identified as a promising candidate for broomrape biocontrol. Phytotoxicity tests of crude extracts from the fungus alone or in interaction with broomrape on P. ramosa microcalli and quantification of necrosis by image analysis confirmed the phytotoxic potential of F. venenatum MIAE02836 metabolites towards the early developmental stages of P. ramosa. Data analysis of a non-targeted metabolomics approach revealed numerous metabolites produced by F. venenatum MIAE02836. Four of them, accumulated during interaction with the parasitic plant, are known for their phytotoxic potential: maculosin, cyclo(Leu-Phe), phenylalanyl-D-histidine and anguidine. These results suggest that combining image acquisition of the microcalli screening test and untargeted metabolomic approach is an interesting and relevant method to characterize phytotoxic fungal metabolites.
Keywords: Fusarium venenatum; Phelipanche ramosa; fungal metabolites; image analysis; phytotoxicity; untargeted metabolomics.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures






References
-
- Gibot-Leclerc S., Dessaint F., Reibel C., Le Corre V. Phelipanche Ramosa (L.) Pomel Populations Differ in Life-History and Infection Response to Hosts. Flora-Morphol. Distrib. Funct. Ecol. Plants. 2013;208:247–252. doi: 10.1016/j.flora.2013.03.007. - DOI
-
- Pointurier O., Gibot-Leclerc S., Moreau D., Reibel C., Vieren E., Colbach N. Designing a Model to Investigate Cropping Systems Aiming to Control Both Parasitic Plants and Weeds. Eur. J. Agron. 2021;129:126318. doi: 10.1016/j.eja.2021.126318. - DOI
-
- Monteiro A., Santos S. Sustainable Approach to Weed Management: The Role of Precision Weed Management. Agronomy. 2022;12:118. doi: 10.3390/agronomy12010118. - DOI
Publication types
MeSH terms
Substances
Grants and funding
- Mycoherbicides for the biocontrol of broomrape, a parasitic plant of numerous crops) project; 2021-2023, grant "Appel à projets scientifiques "innovation" IB 2021"/Institut National de Recherche pour l'Agriculture, l'Alimentation et l'Environnement
- Formation complémentaire par la recherche/French Ministry of Agriculture and Food
LinkOut - more resources
Full Text Sources
