

The PA-X host shutoff site 100 V exerts a contrary effect on viral fitness of the highly pathogenic H7N9 influenza A virus in mice and chickens
- PMID: 39731774
- PMCID: PMC11702944
- DOI: 10.1080/21505594.2024.2445238
The PA-X host shutoff site 100 V exerts a contrary effect on viral fitness of the highly pathogenic H7N9 influenza A virus in mice and chickens
Abstract
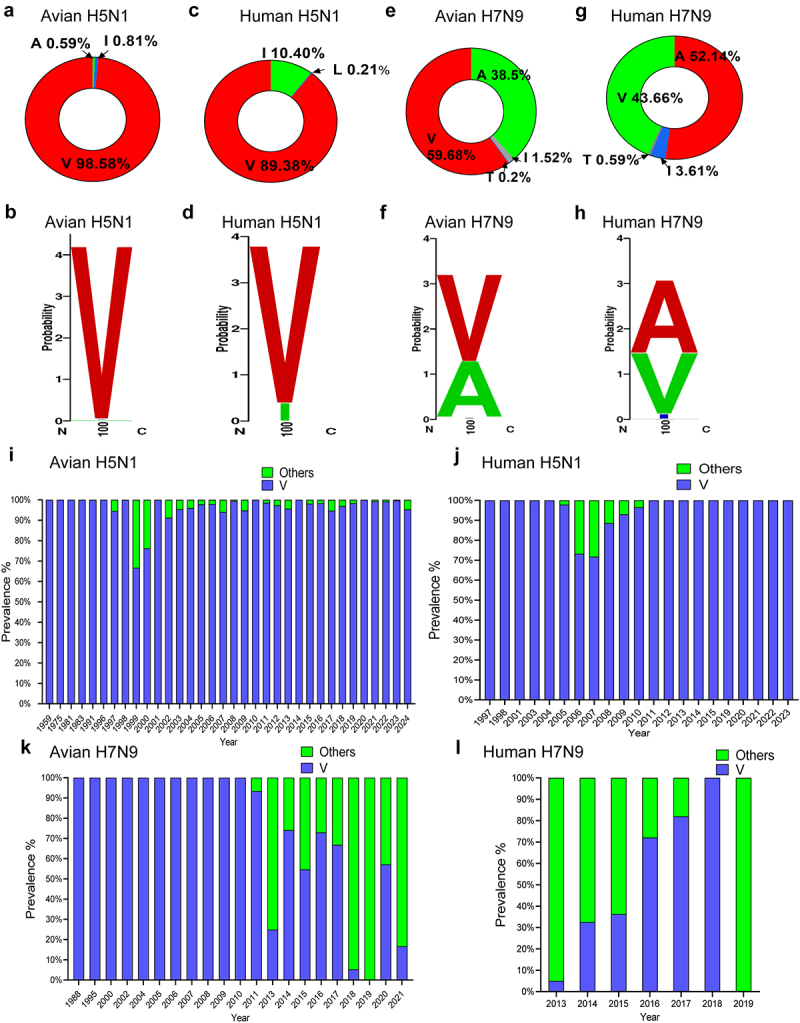
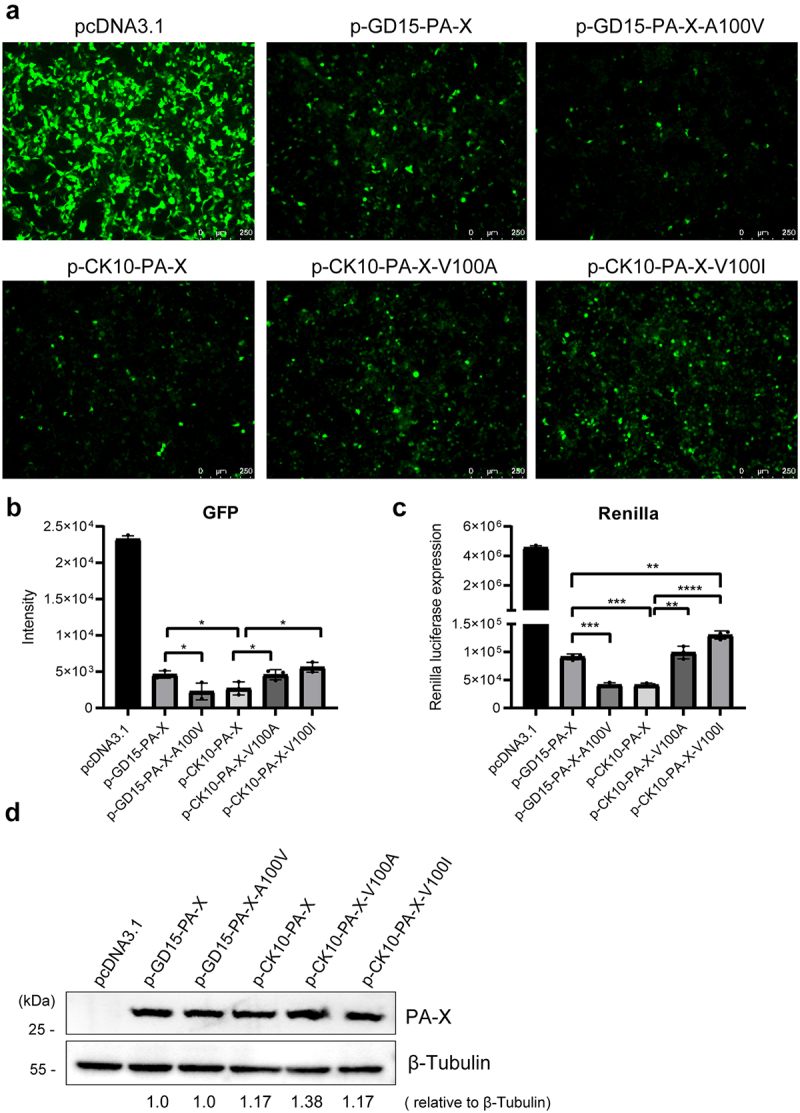
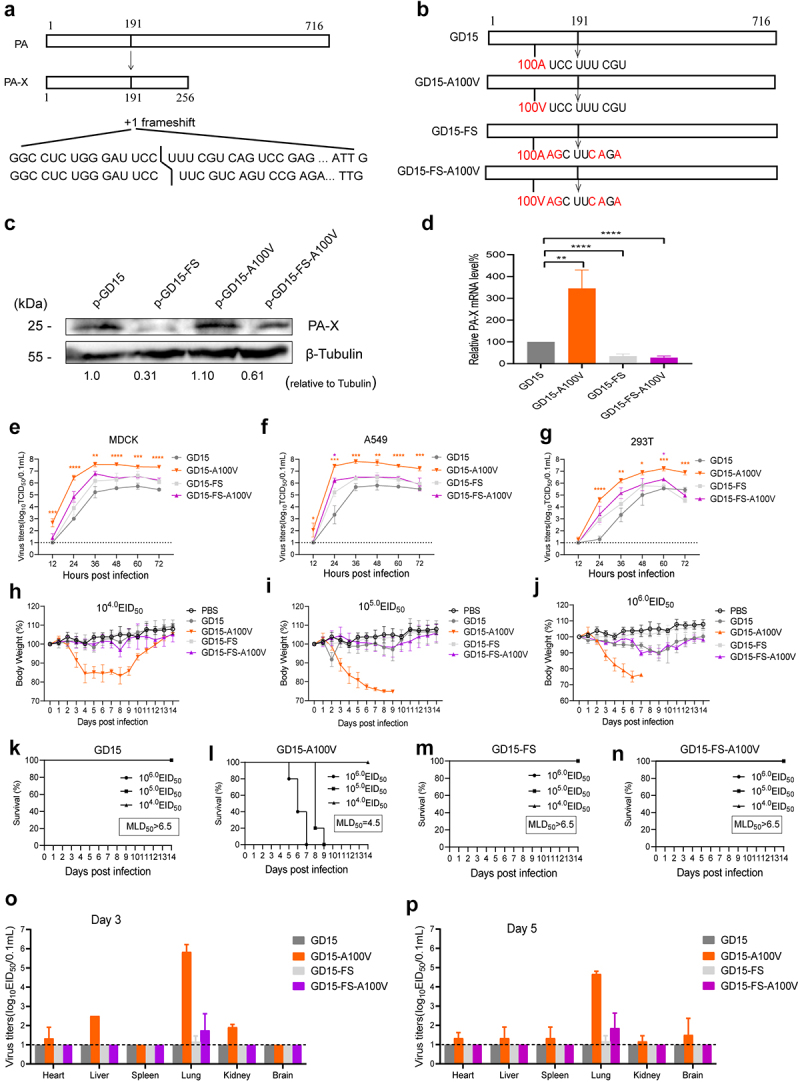
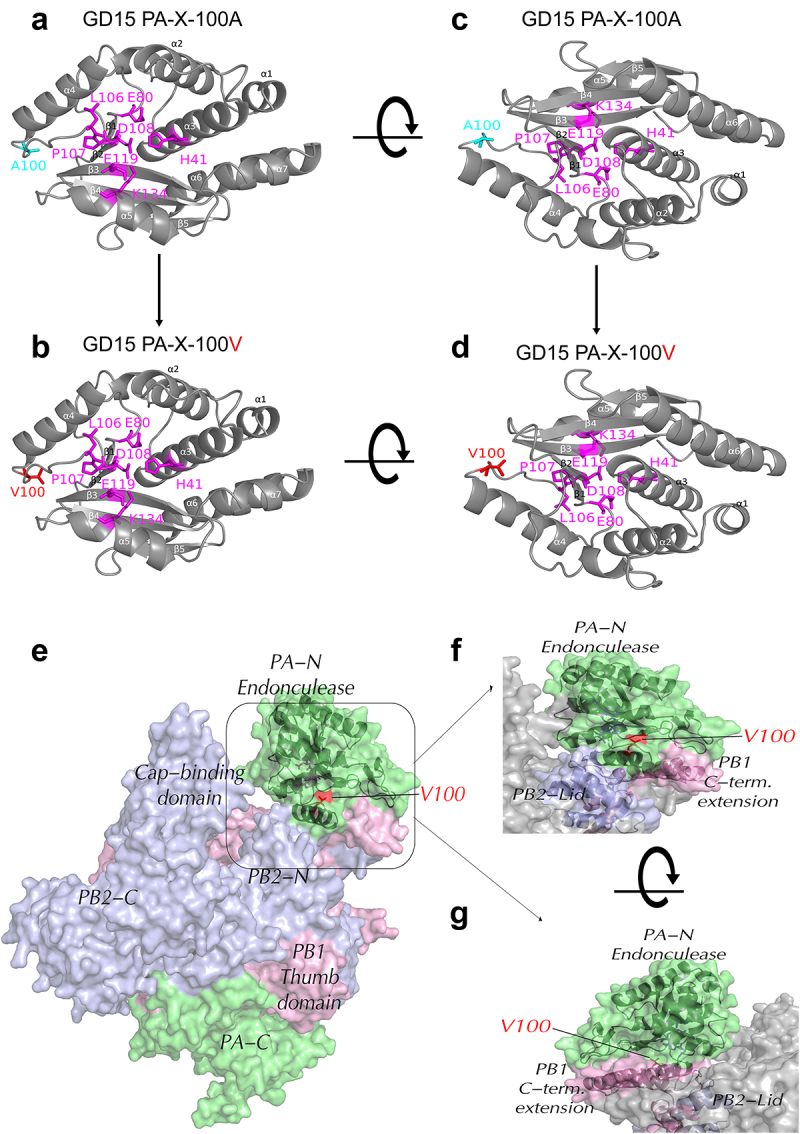
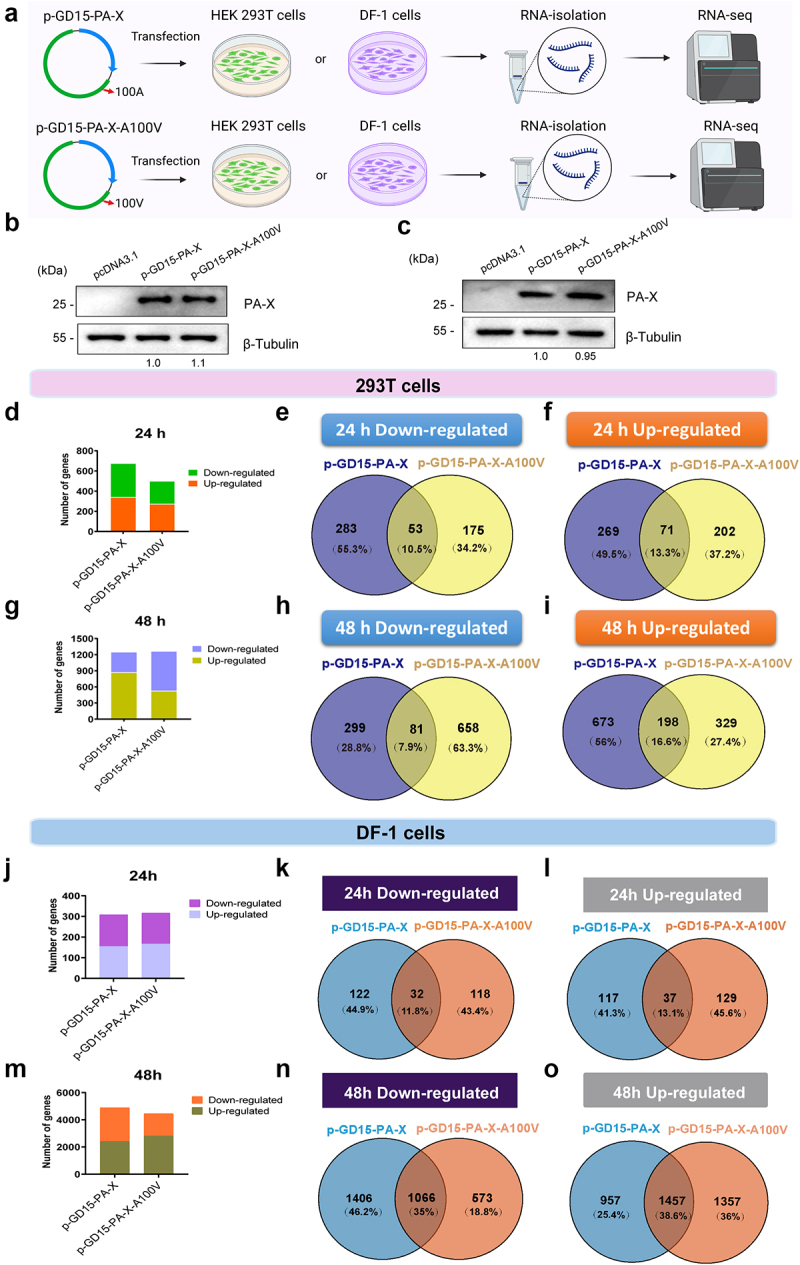
Several viruses, including influenza A virus (IAV), encode viral factors to hijack cellular RNA biogenesis processes to direct the degradation of host mRNAs, termed "host shutoff." Host shutoff enables viruses to simultaneously reduce antiviral responses and provides preferential access for viral mRNAs to cellular translation machinery. IAV PA-X is one of these factors that selectively shuts off the global host genes. However, the specific role of PA-X host shutoff activity in viral fitness of IAV remains poorly understood. Herein, we successfully mapped PA-X 100 V as a novel site important for host shutoff of the H7N9 and H5N1 viruses. By analysing the polymorphism of this residue in various subtype viruses, we found that PA-X 100 was highly variable in H7N9 viruses. Structural analysis revealed that 100 V was generally close to the PA-X endonuclease active site, which may account for its host shutoff activity. By generating the corresponding mutant viruses derived from the parental H7N9 virus and the PA-X-deficient H7N9 virus, we determined that PA-X 100 V significantly enhanced viral fitness in mice while diminishing viral virulence in chickens. Mechanistically, PA-X 100 V significantly increased viral polymerase activity and viral replication in mammalian cells. Furthermore, PA-X 100 V highly blunted the global host response in 293T cells, particularly restraining genes involved in energy metabolism and inflammatory response. Collectively, our data provided information about the intricate role of the PA-X host shutoff site in regulating the viral fitness of the H7N9 influenza virus, which furthers our understanding of the complicated pathogenesis of the influenza A virus.
Keywords: H7N9 avian influenza virus; PA-X; chickens; host shutoff; mice; viral fitness.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
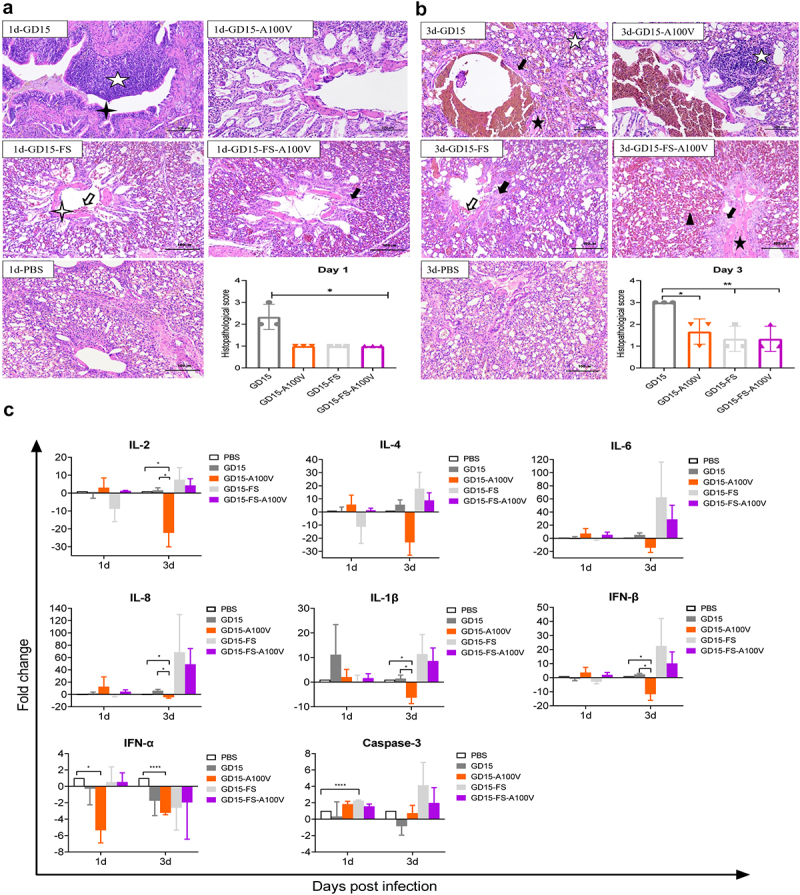
 , lung congestion and haemorrhage;
, lung congestion and haemorrhage;  , necrotic detached cells were seen in the bronchiolar lumen;
, necrotic detached cells were seen in the bronchiolar lumen;  , lymphocyte infiltration in pulmonary alveoli bronchiolar lumen and around blood vessel. (c) Innate immune response in mice. Groups of mice were infected with each of the indicated recombinant virus at a dose of 106.0 EID50. Three mice of each group were euthanized on day 3 and 5 p.t. For determination of cytokine response in mouse lung. The concentration of cytokines/chemokines and complements derived components in mouse lung was analysed by ELISA. Values were shown as the means ± SD of three samples.
, lymphocyte infiltration in pulmonary alveoli bronchiolar lumen and around blood vessel. (c) Innate immune response in mice. Groups of mice were infected with each of the indicated recombinant virus at a dose of 106.0 EID50. Three mice of each group were euthanized on day 3 and 5 p.t. For determination of cytokine response in mouse lung. The concentration of cytokines/chemokines and complements derived components in mouse lung was analysed by ELISA. Values were shown as the means ± SD of three samples.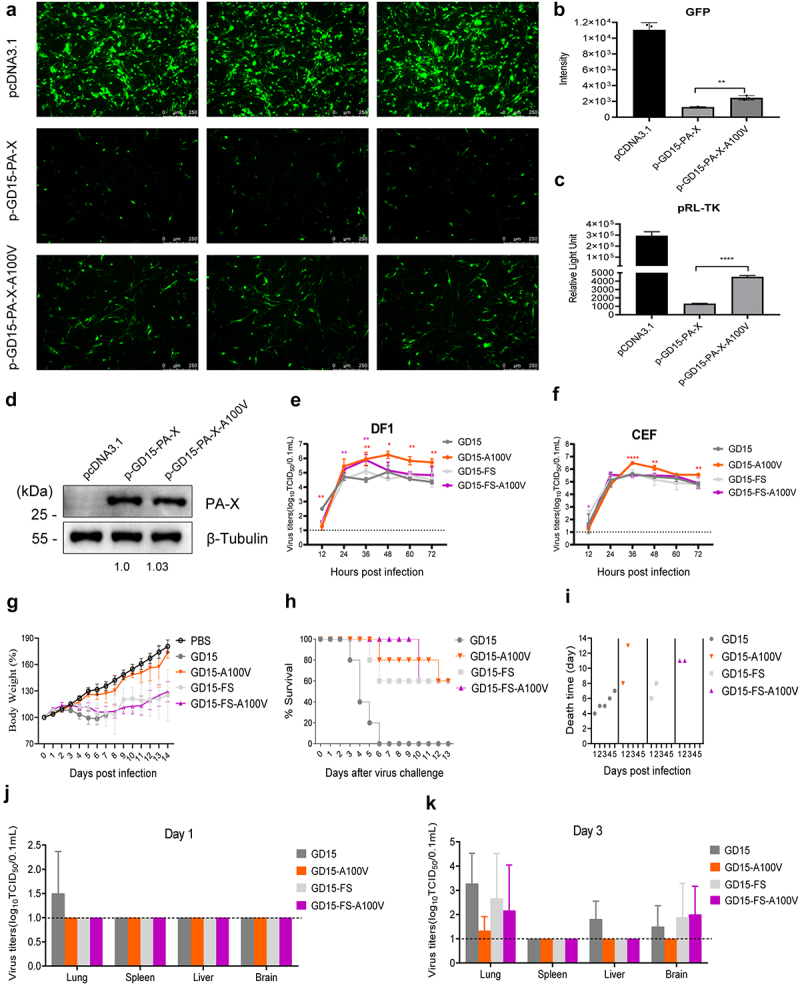
 , lung haemorrhage;
, lung haemorrhage;  , lung congestion;
, lung congestion;  , dilation of parabrochus and structure disappearance of the lung chamber;
, dilation of parabrochus and structure disappearance of the lung chamber;  , lymphocyte infiltration in blood vessels or parabrochus and/or pulmonary chamber;
, lymphocyte infiltration in blood vessels or parabrochus and/or pulmonary chamber;  , eosinophilic material can be seen in the cavity;
, eosinophilic material can be seen in the cavity;  , the structure of the exhale capillary was unclear;
, the structure of the exhale capillary was unclear;  , water degeneration, cell swelling and cytoplasm light dye of the epithelial cells in bronchus;
, water degeneration, cell swelling and cytoplasm light dye of the epithelial cells in bronchus;  , a small amount of epithelial cells was lost in the accessory bronchus. (c) Groups of chickens were infected with each of the indicated recombinant virus at a dose of 106.0 EID50. Three chickens of each group were euthanized on day 1 and 3 p.t. for the determination of cytokine response in chicken lungs. The concentration of cytokine/chemokine and complement-derived components in chicken lungs was analysed by qRT-PCR. Values were shown as the means ± SD of three samples.
, a small amount of epithelial cells was lost in the accessory bronchus. (c) Groups of chickens were infected with each of the indicated recombinant virus at a dose of 106.0 EID50. Three chickens of each group were euthanized on day 1 and 3 p.t. for the determination of cytokine response in chicken lungs. The concentration of cytokine/chemokine and complement-derived components in chicken lungs was analysed by qRT-PCR. Values were shown as the means ± SD of three samples.
References
Publication types
MeSH terms
Substances
Associated data
- figshare/10.6084/m9.figshare.26860537
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical