

Social context modulates active avoidance: Contributions of the anterior cingulate cortex in male and female rats
- PMID: 39737250
- PMCID: PMC11683269
- DOI: 10.1016/j.ynstr.2024.100702
Social context modulates active avoidance: Contributions of the anterior cingulate cortex in male and female rats
Abstract
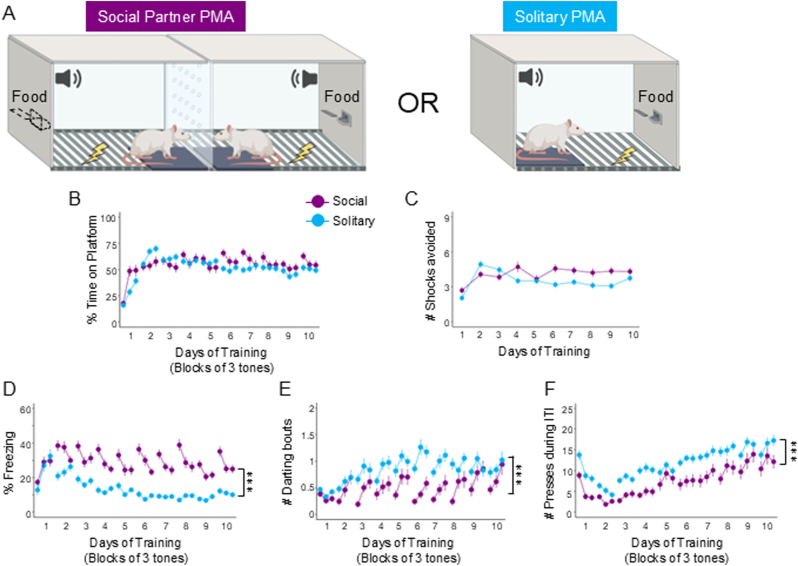
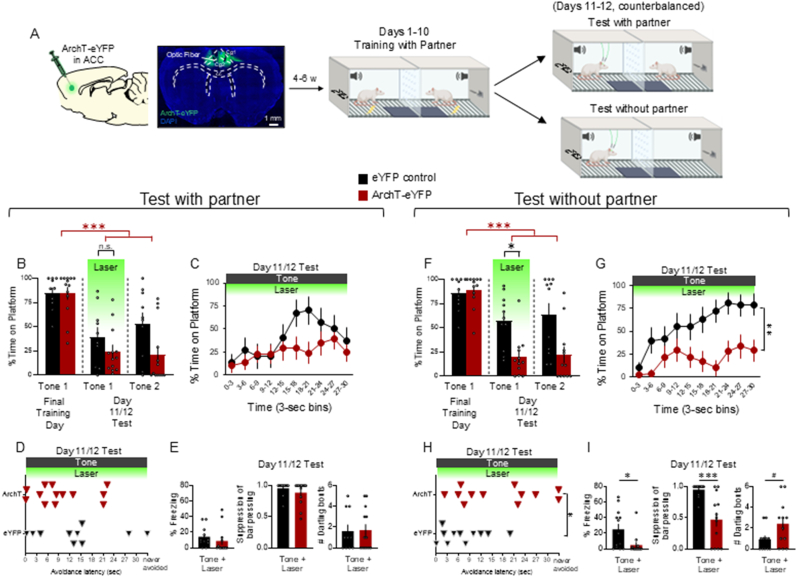
Actively avoiding danger is necessary for survival. Most research on active avoidance has focused on the behavioral and neurobiological processes when individuals learn to avoid alone, within a solitary context. Therefore, little is known about how social context affects active avoidance. Using a modified version of the platform-mediated avoidance task in rats, we investigated whether the presence of a social partner attenuates conditioned freezing and enhances avoidance compared to avoidance in a solitary context. Rats spent a similar amount of time avoiding during either context; however, rats trained in the social context exhibited greater freezing as well as lower rates of darting and food seeking compared to rats trained in the solitary context. In addition, we observed higher levels of avoidance in females compared to males in the solitary context, but this sex difference was not present in rats trained in the social context. To gain greater mechanistic insight, we optogenetically inactivated glutamatergic projection neurons in the anterior cingulate cortex (ACC) following avoidance training in either context. After avoidance was learned in a social context, photoinactivation of ACC reduced expression of avoidance during a test when the social partner was absent, but not when the partner was present. Our findings suggest a novel contribution of the ACC in avoidance that is learned with a social partner, which has translational implications for understanding ACC dysfunction in those suffering from trauma-related disorders.
Keywords: Fear; Optogenetics; Prefrontal cortex; Sex differences.
© 2024 The Authors.
Conflict of interest statement
None.
Figures


Update of
-
Active avoidance recruits the anterior cingulate cortex regardless of social context in male and female rats.Res Sq [Preprint]. 2024 Jun 7:rs.3.rs-3750422. doi: 10.21203/rs.3.rs-3750422/v2. Res Sq. 2024. Update in: Neurobiol Stress. 2024 Dec 05;34:100702. doi: 10.1016/j.ynstr.2024.100702. PMID: 38260416 Free PMC article. Updated. Preprint.
Similar articles
-
Modifying the platform-mediated avoidance task: A new protocol to study active avoidance within a social context in rats.PLoS One. 2025 Apr 29;20(4):e0321776. doi: 10.1371/journal.pone.0321776. eCollection 2025. PLoS One. 2025. PMID: 40299933 Free PMC article.
-
Active avoidance recruits the anterior cingulate cortex regardless of social context in male and female rats.Res Sq [Preprint]. 2024 Jun 7:rs.3.rs-3750422. doi: 10.21203/rs.3.rs-3750422/v2. Res Sq. 2024. Update in: Neurobiol Stress. 2024 Dec 05;34:100702. doi: 10.1016/j.ynstr.2024.100702. PMID: 38260416 Free PMC article. Updated. Preprint.
-
Adolescent alcohol exposure alters threat avoidance in adulthood.Front Behav Neurosci. 2023 Jan 25;16:1098343. doi: 10.3389/fnbeh.2022.1098343. eCollection 2022. Front Behav Neurosci. 2023. PMID: 36761697 Free PMC article.
-
Acute stress yields a sex-dependent facilitation of signaled active avoidance in rats.Neurobiol Stress. 2024 Jun 13;31:100656. doi: 10.1016/j.ynstr.2024.100656. eCollection 2024 Jul. Neurobiol Stress. 2024. PMID: 38994219 Free PMC article. Review.
-
Conserved features of anterior cingulate networks support observational learning across species.Neurosci Biobehav Rev. 2019 Dec;107:215-228. doi: 10.1016/j.neubiorev.2019.09.009. Epub 2019 Sep 8. Neurosci Biobehav Rev. 2019. PMID: 31509768 Free PMC article. Review.
Cited by
-
Behavioral outputs and overlapping circuits between conditional fear and active avoidance.Neurobiol Learn Mem. 2024 Sep;213:107943. doi: 10.1016/j.nlm.2024.107943. Epub 2024 May 29. Neurobiol Learn Mem. 2024. PMID: 38821256 Free PMC article. Review.
-
Modifying the platform-mediated avoidance task: A new protocol to study active avoidance within a social context in rats.PLoS One. 2025 Apr 29;20(4):e0321776. doi: 10.1371/journal.pone.0321776. eCollection 2025. PLoS One. 2025. PMID: 40299933 Free PMC article.
-
Dopaminergic projections to the prefrontal cortex are critical for rapid threat avoidance learning.Curr Biol. 2025 Sep 8;35(17):4259-4269.e3. doi: 10.1016/j.cub.2025.07.035. Epub 2025 Aug 8. Curr Biol. 2025. PMID: 40782805
References
-
- American Psychiatric Association . fifth ed. American Psychiatric Association; 2013. Diagnostic and Statistical Manual of Mental Disorders : DSM-5.
Grants and funding
LinkOut - more resources
Full Text Sources
