

Rapid nongenomic estrogen signaling controls alcohol drinking behavior in mice
- PMID: 39737915
- PMCID: PMC11686278
- DOI: 10.1038/s41467-024-54737-6
Rapid nongenomic estrogen signaling controls alcohol drinking behavior in mice
Abstract
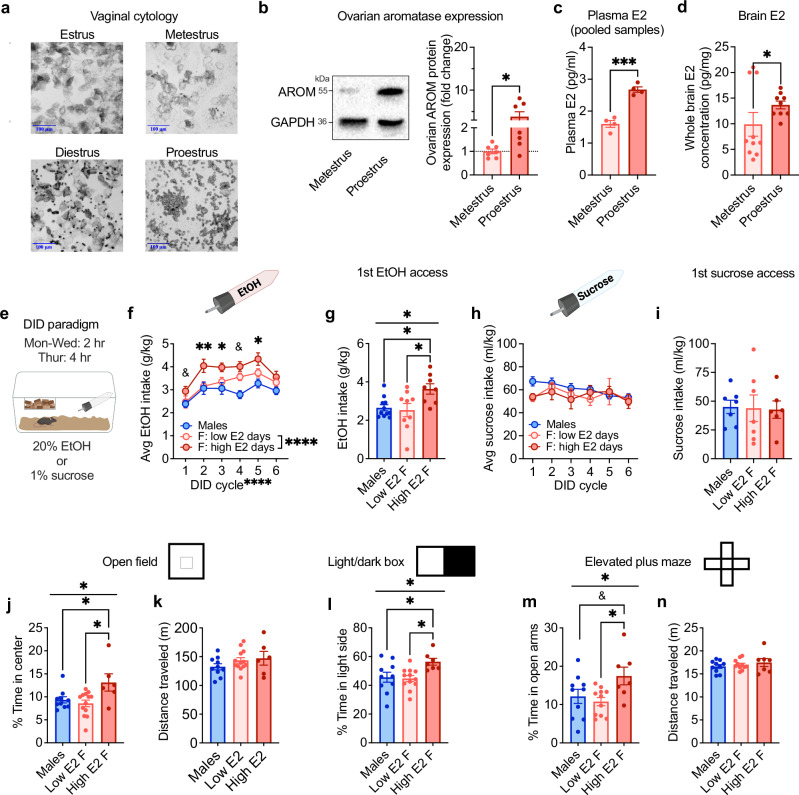
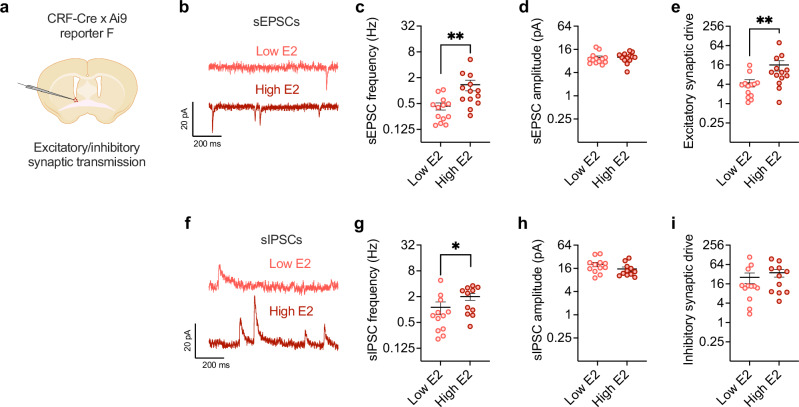
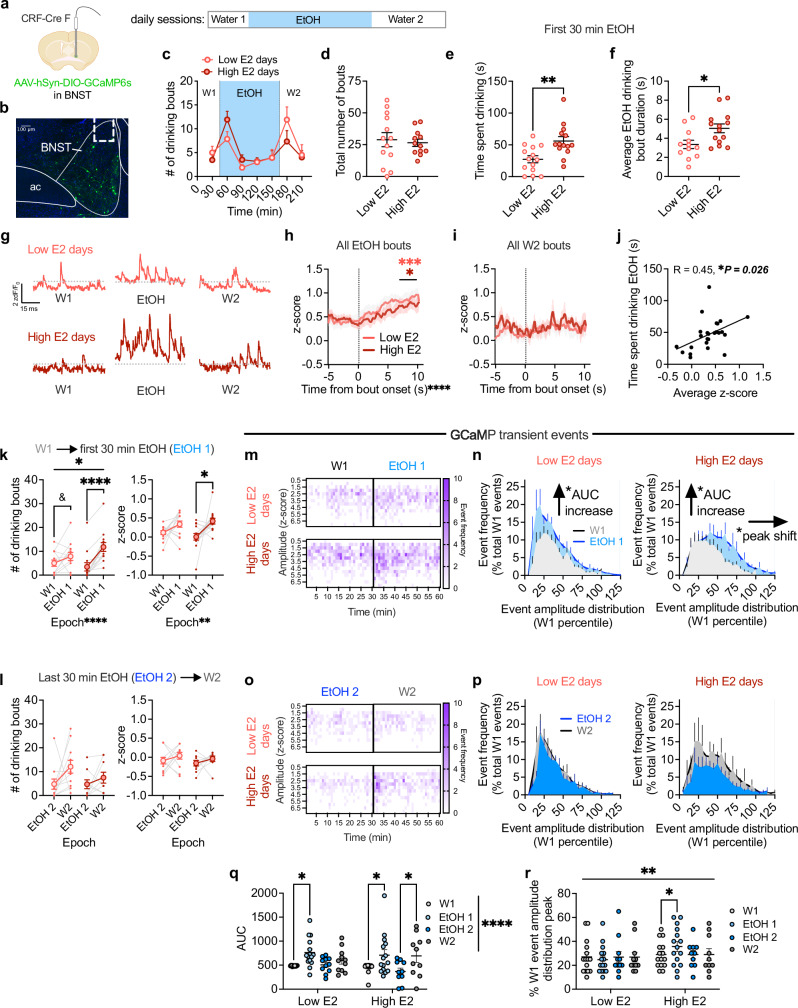
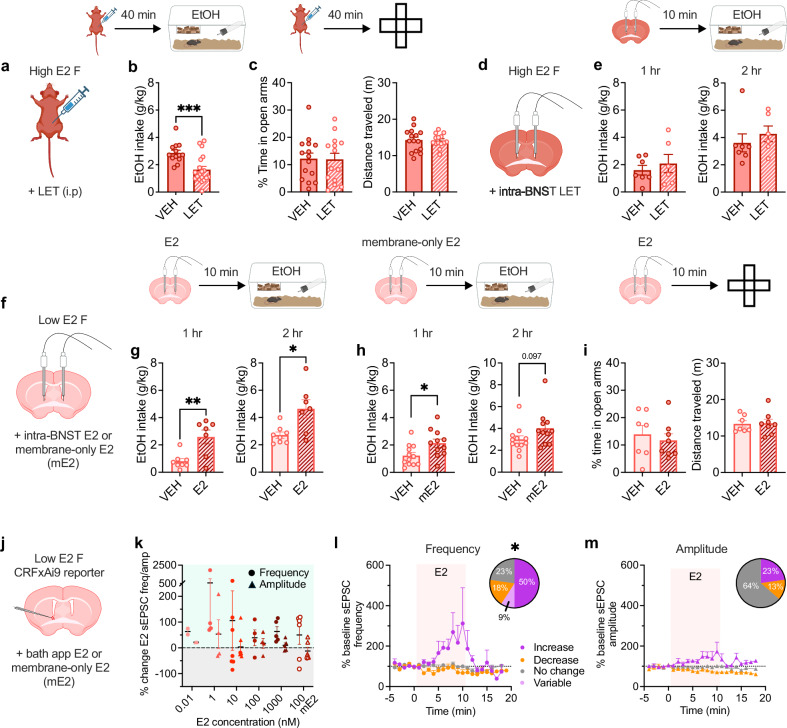
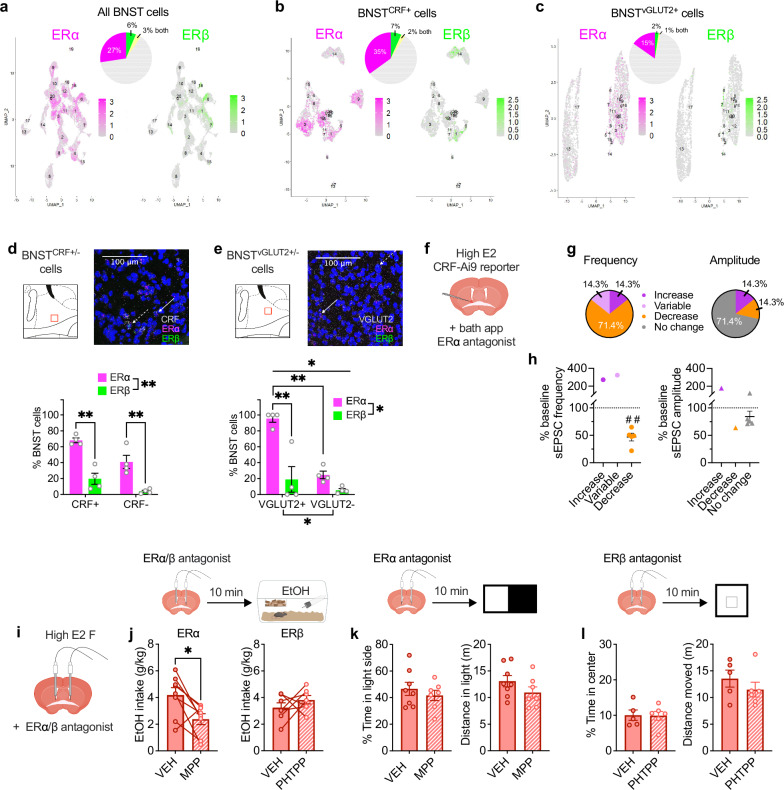
Ovarian-derived estrogen can signal non-canonically at membrane-associated receptors in the brain to rapidly regulate neuronal function. Early alcohol drinking confers greater risk for alcohol use disorder in women than men, and binge alcohol drinking is correlated with high estrogen levels, but a causal role for estrogen in driving alcohol drinking has not been established. We found that female mice displayed greater binge alcohol drinking and reduced avoidance when estrogen was high during the estrous cycle than when it was low. The pro-drinking, but not anxiolytic, effect of high endogenous estrogen occurred via rapid signaling at membrane-associated estrogen receptor alpha in the bed nucleus of the stria terminalis, which promoted synaptic excitation of corticotropin-releasing factor neurons and facilitated their activity during alcohol drinking. Thus, this study demonstrates a rapid, nongenomic signaling mechanism for ovarian-derived estrogen in the brain controlling behavior in gonadally intact females.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
Update of
-
Rapid nongenomic estrogen signaling controls alcohol drinking behavior.bioRxiv [Preprint]. 2024 Aug 15:2023.11.02.565358. doi: 10.1101/2023.11.02.565358. bioRxiv. 2024. Update in: Nat Commun. 2024 Dec 30;15(1):10725. doi: 10.1038/s41467-024-54737-6. PMID: 37961707 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 AA027645/AA/NIAAA NIH HHS/United States
- T32 GM141949/GM/NIGMS NIH HHS/United States
- 516215326/Deutsche Forschungsgemeinschaft (German Research Foundation)
- DA039080/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- T32GM007739/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- T32 GM007739/GM/NIGMS NIH HHS/United States
- AA023559/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R00 AA023559/AA/NIAAA NIH HHS/United States
- P51 OD011092/OD/NIH HHS/United States
- AA027645/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- F31 AA029293/AA/NIAAA NIH HHS/United States
- AA029293/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- OD011092/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- T32 DA039080/DA/NIDA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
