

Malate initiates a proton-sensing pathway essential for pH regulation of inflammation
- PMID: 39737965
- PMCID: PMC11683149
- DOI: 10.1038/s41392-024-02076-9
Malate initiates a proton-sensing pathway essential for pH regulation of inflammation
Abstract
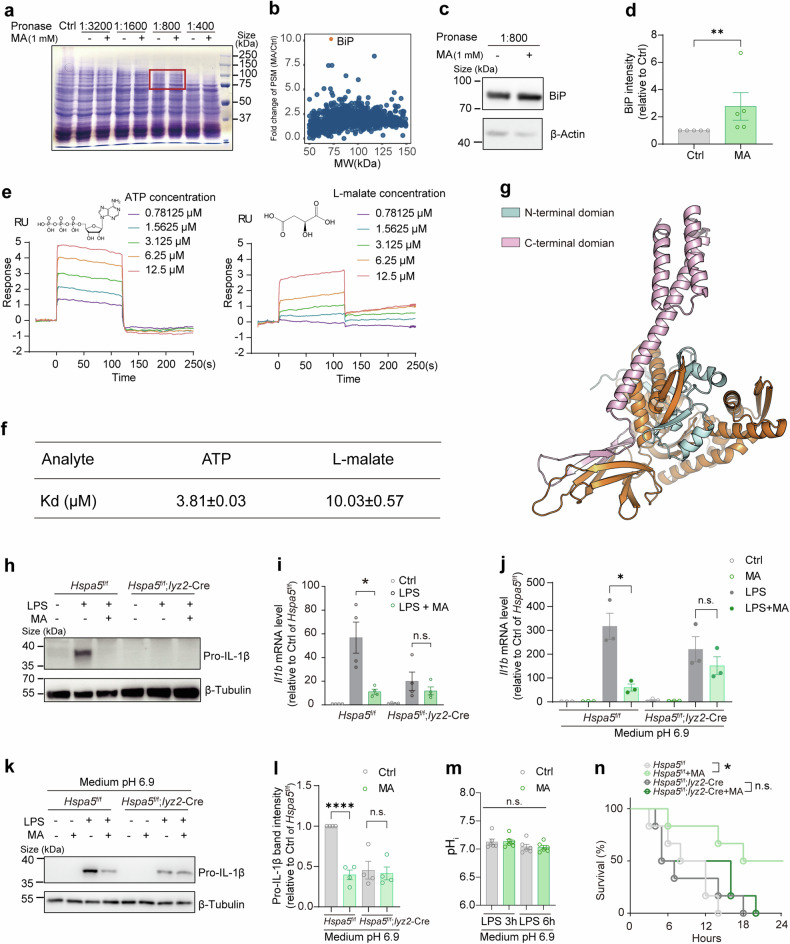
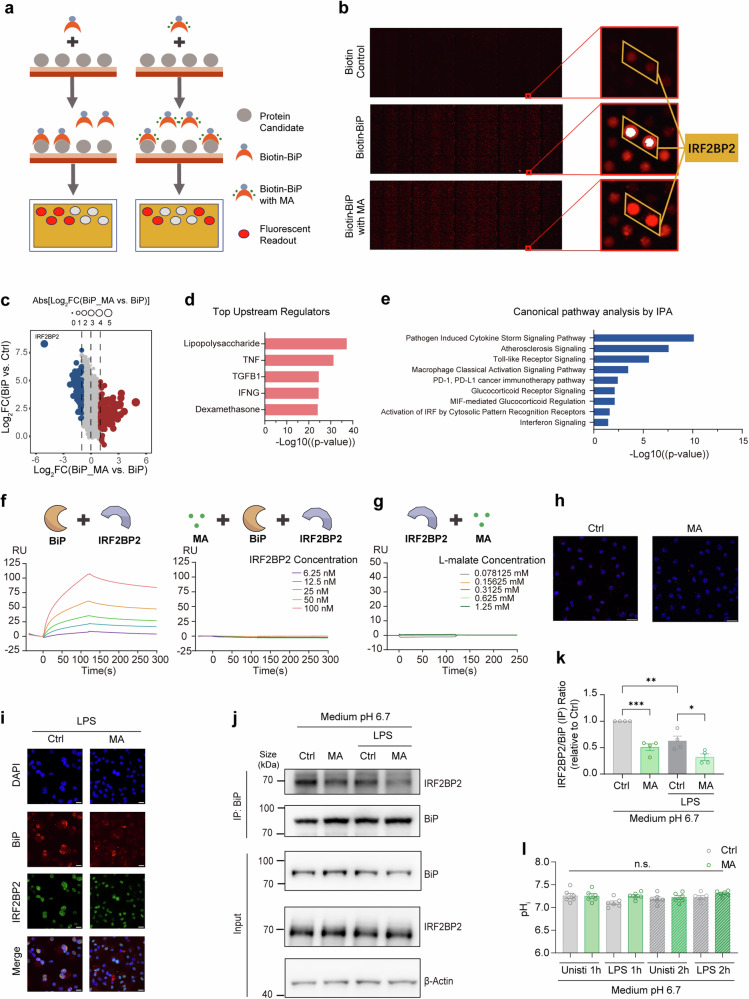
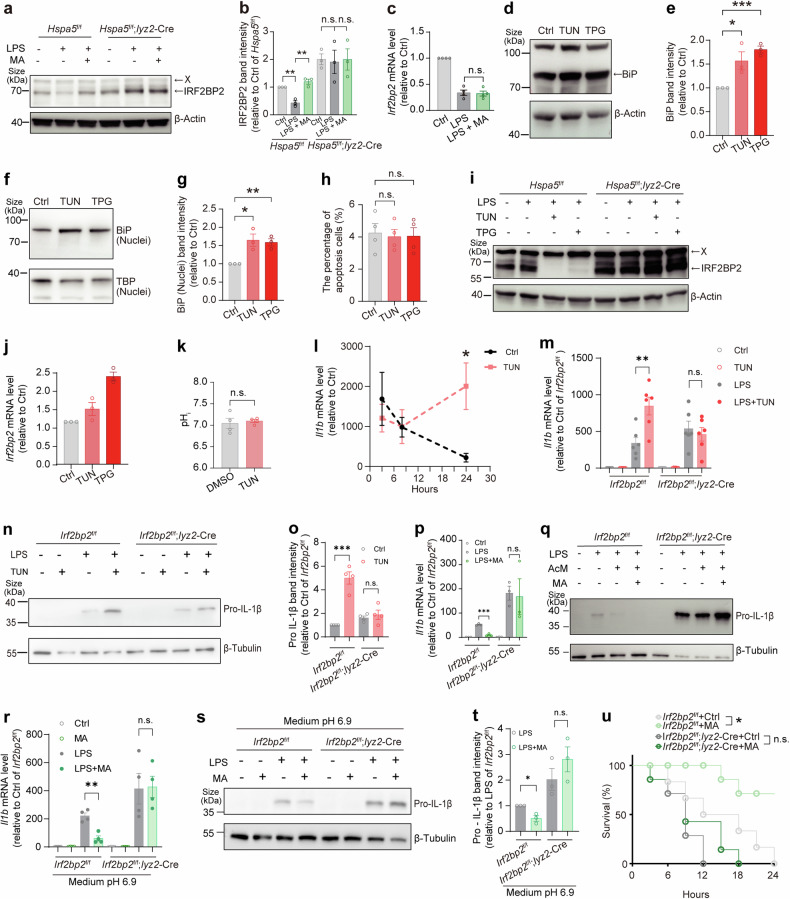
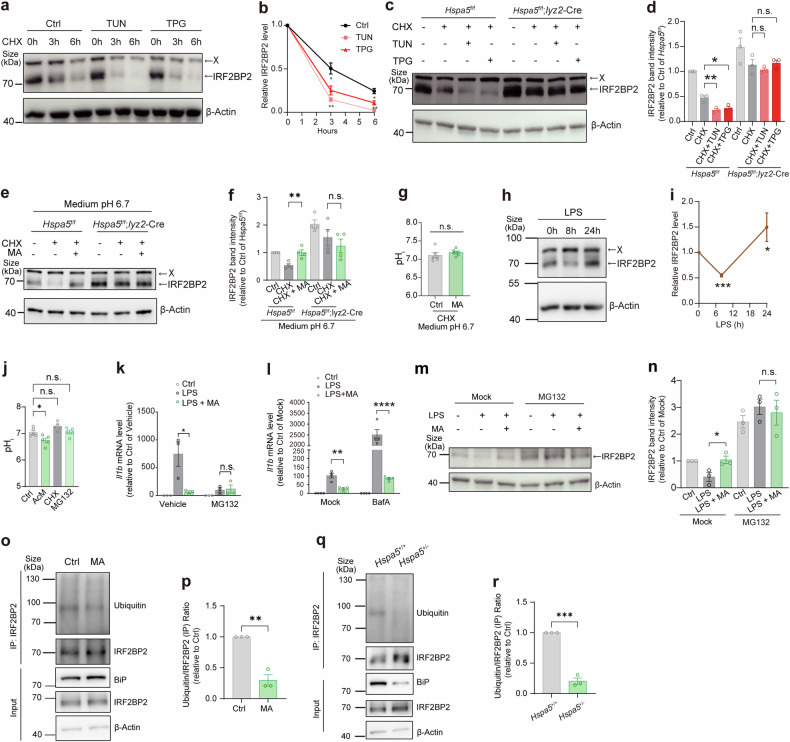
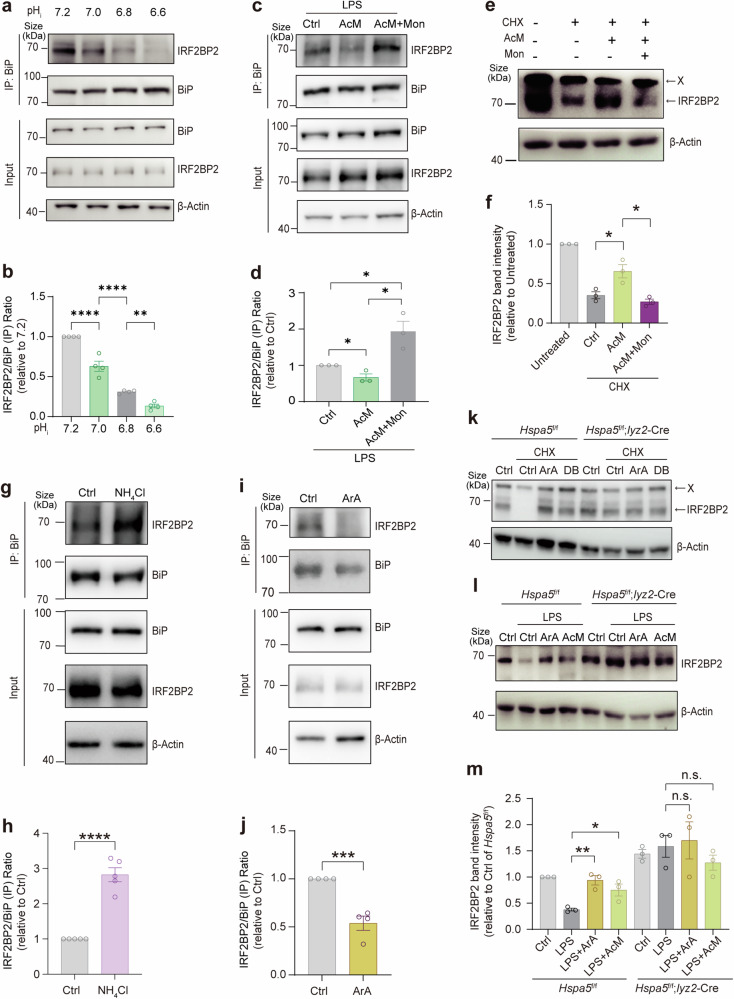
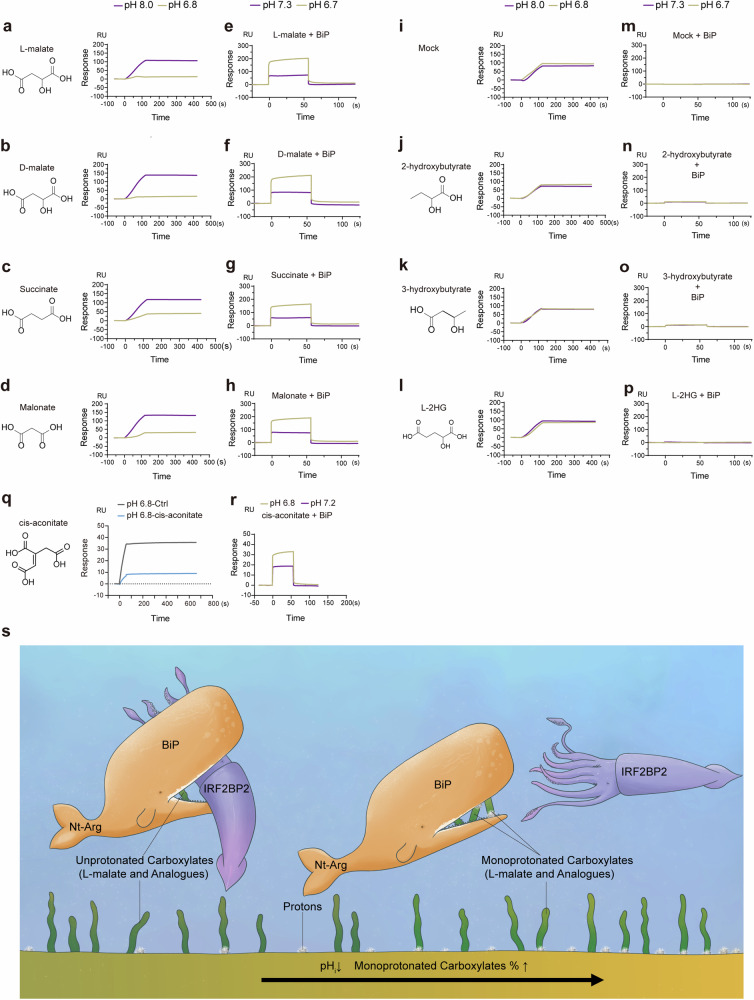
Metabolites can double as a signaling modality that initiates physiological adaptations. Metabolism, a chemical language encoding biological information, has been recognized as a powerful principle directing inflammatory responses. Cytosolic pH is a regulator of inflammatory response in macrophages. Here, we found that L-malate exerts anti-inflammatory effect via BiP-IRF2BP2 signaling, which is a sensor of cytosolic pH in macrophages. First, L-malate, a TCA intermediate upregulated in pro-inflammatory macrophages, was identified as a potent anti-inflammatory metabolite through initial screening. Subsequent screening with DARTS and MS led to the isolation of L-malate-BiP binding. Further screening through protein‒protein interaction microarrays identified a L-malate-restrained coupling of BiP with IRF2BP2, a known anti-inflammatory protein. Interestingly, pH reduction, which promotes carboxyl protonation of L-malate, facilitates L-malate and carboxylate analogues such as succinate to bind BiP, and disrupt BiP-IRF2BP2 interaction in a carboxyl-dependent manner. Both L-malate and acidification inhibit BiP-IRF2BP2 interaction, and protect IRF2BP2 from BiP-driven degradation in macrophages. Furthermore, both in vitro and in vivo, BiP-IRF2BP2 signal is required for effects of both L-malate and pH on inflammatory responses. These findings reveal a previously unrecognized, proton/carboxylate dual sensing pathway wherein pH and L-malate regulate inflammatory responses, indicating the role of certain carboxylate metabolites as adaptors in the proton biosensing by interactions between macromolecules.
© 2024. The Author(s).
Conflict of interest statement
Conflict of interest: Hongming Miao is the editorial board member of Signal Transduction and Targeted Therapy, but he has not been involved in the process of the manuscript handling. Other authors declare no competing interest.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
