

Cerebellar-driven cortical dynamics can enable task acquisition, switching and consolidation
- PMID: 39738061
- PMCID: PMC11686095
- DOI: 10.1038/s41467-024-55315-6
Cerebellar-driven cortical dynamics can enable task acquisition, switching and consolidation
Abstract
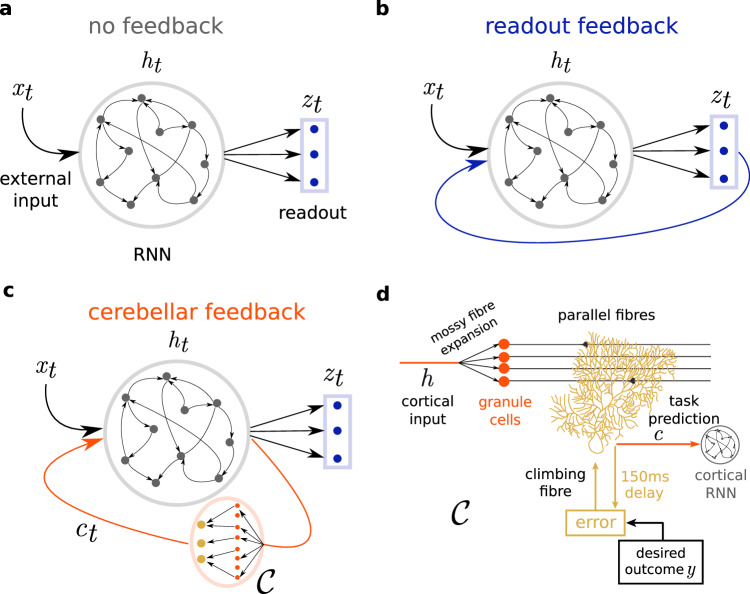
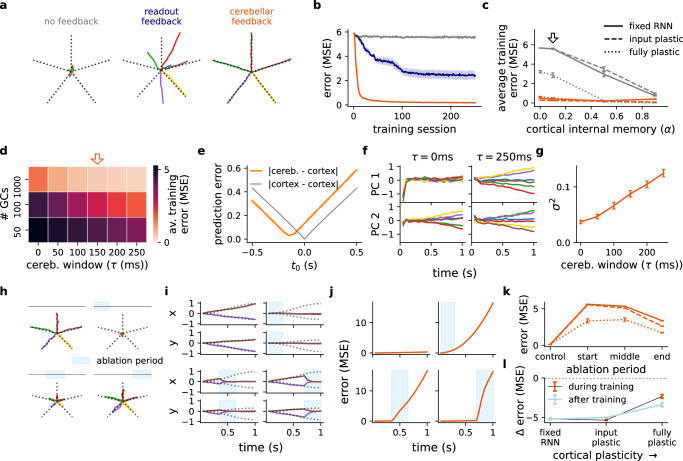
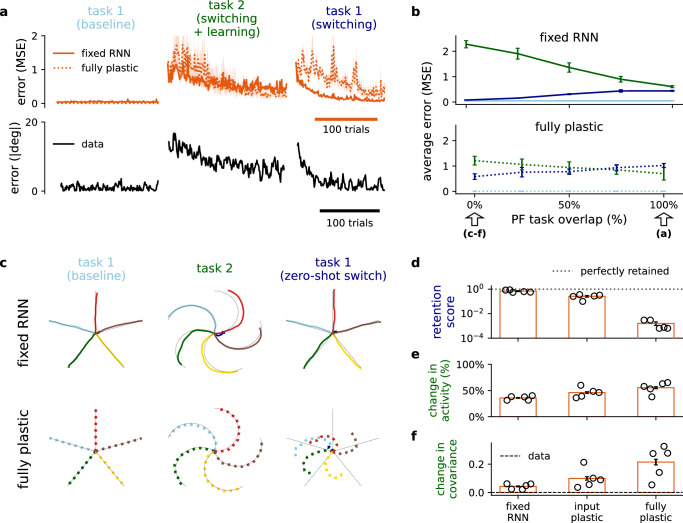
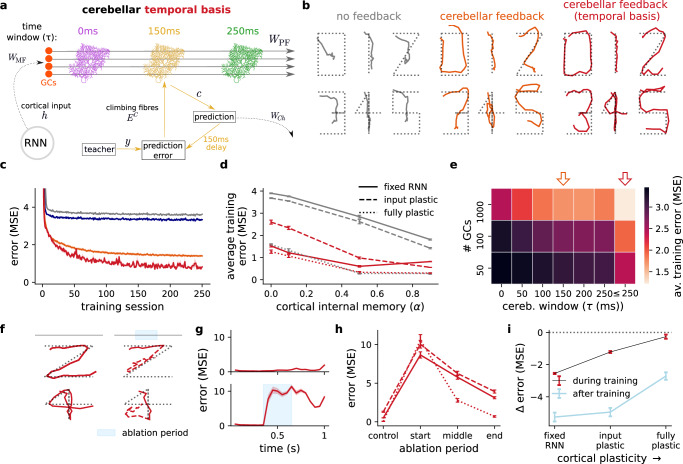
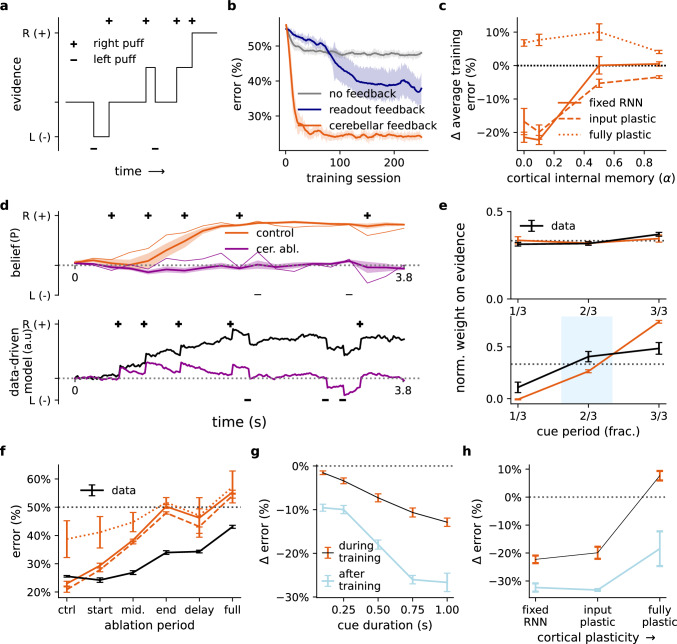
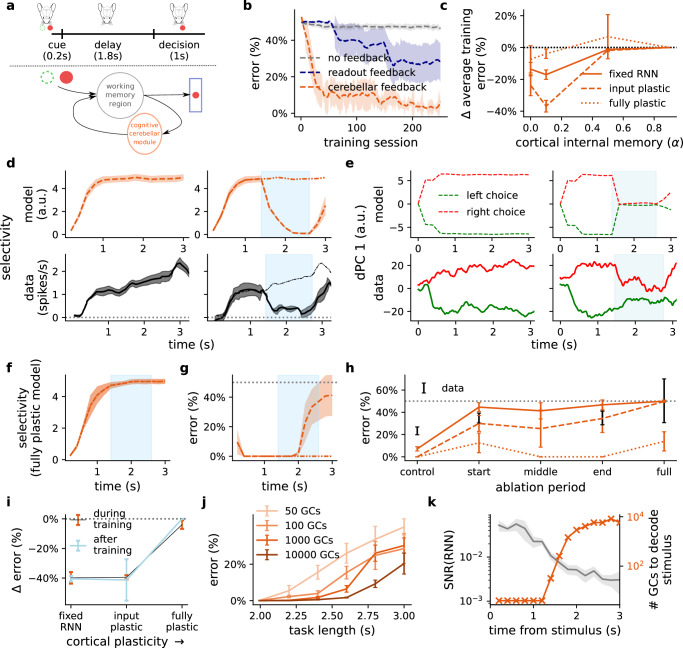
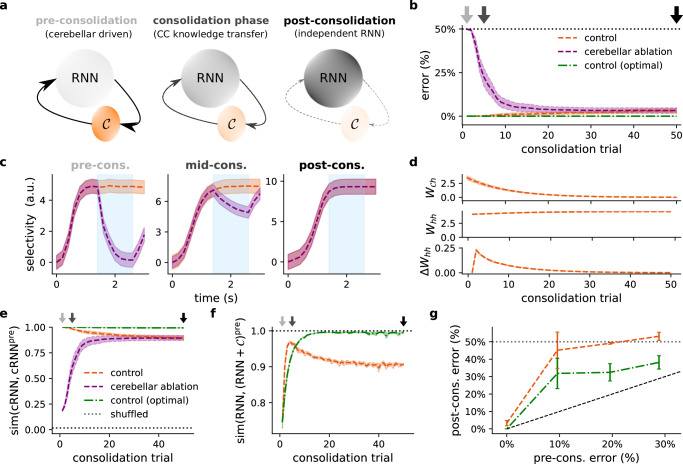
The brain must maintain a stable world model while rapidly adapting to the environment, but the underlying mechanisms are not known. Here, we posit that cortico-cerebellar loops play a key role in this process. We introduce a computational model of cerebellar networks that learn to drive cortical networks with task-outcome predictions. First, using sensorimotor tasks, we show that cerebellar feedback in the presence of stable cortical networks is sufficient for rapid task acquisition and switching. Next, we demonstrate that, when trained in working memory tasks, the cerebellum can also underlie the maintenance of cognitive-specific dynamics in the cortex, explaining a range of optogenetic and behavioural observations. Finally, using our model, we introduce a systems consolidation theory in which task information is gradually transferred from the cerebellum to the cortex. In summary, our findings suggest that cortico-cerebellar loops are an important component of task acquisition, switching, and consolidation in the brain.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
- EP/R513179/1/RCUK | Engineering and Physical Sciences Research Council (EPSRC)
- EP/X029336/1/RCUK | Engineering and Physical Sciences Research Council (EPSRC)
- 209453/Z/17/Z/Wellcome Trust (Wellcome)
- MR/X006107/1/RCUK | Medical Research Council (MRC)
- BB/X013340/1/RCUK | Biotechnology and Biological Sciences Research Council (BBSRC)
LinkOut - more resources
Full Text Sources
