

Apex1 safeguards genomic stability to ensure a cytopathic T cell fate in autoimmune disease models
- PMID: 39739423
- PMCID: PMC11827838
- DOI: 10.1172/JCI183671
Apex1 safeguards genomic stability to ensure a cytopathic T cell fate in autoimmune disease models
Abstract
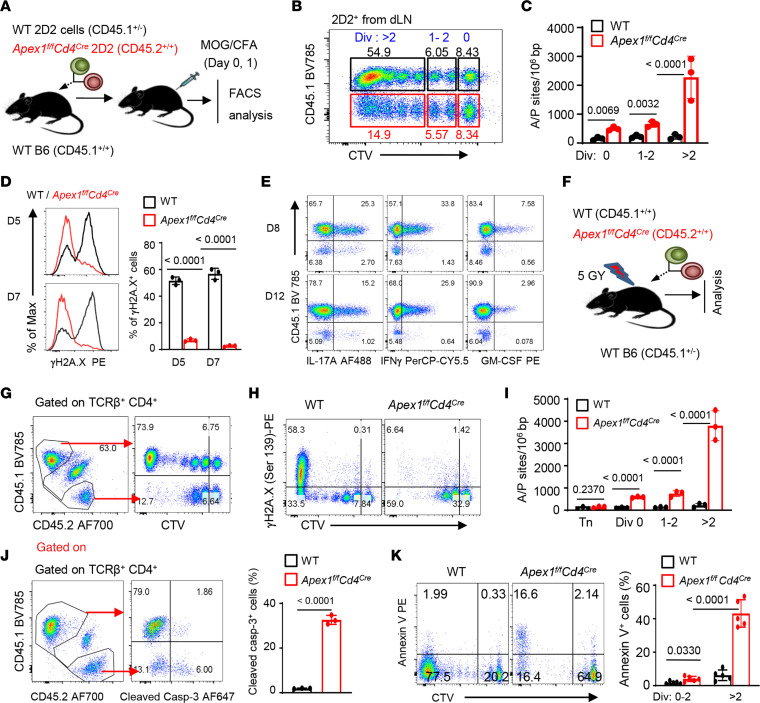
T cells have a remarkable capacity to clonally expand, a process that is intricately linked to their effector activities. As vigorously proliferating T cell also incur substantial DNA lesions, how the dividing T cells safeguard their genomic integrity to allow the generation of T effector cells remains largely unknown. Here we report the identification of the apurinic/apyrimidinic endonuclease-1 (Apex1) as an indispensable molecule for the induction of cytopathic T effectors in mouse models. We demonstrate that conditional deletion of Apex1 in T cells resulted in a remarkable accumulation of baseless DNA sites in the genome of proliferating T cells, which further led to genomic instability and apoptotic cell death. Consequently, Apex1-deleted T cells failed to acquire any effector features after activation and failed to mediate autoimmune diseases and allergic tissue damages. Detailed mutational analyses pinpointed the importance of its endonuclease domain in the generation of T effector cells. We provide further evidence that inhibiting the base repair activities of Apex1 with chemical inhibitors similarly abrogated the induction of autoimmune diseases. Collectively, our study suggests that Apex1 serves as a gatekeeper for the generation of cytopathic T cells and that therapeutically targeting Apex1 may have important clinical implications in the treatment of autoimmune diseases.
Keywords: Autoimmune diseases; Autoimmunity; Immunology; T cells.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
