

A scaleable inducible knockout system for studying essential gene function in the malaria parasite
- PMID: 39739757
- PMCID: PMC11879119
- DOI: 10.1093/nar/gkae1274
A scaleable inducible knockout system for studying essential gene function in the malaria parasite
Abstract
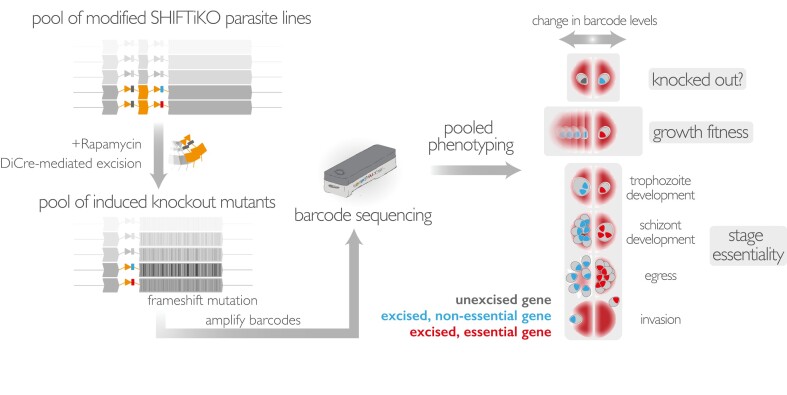
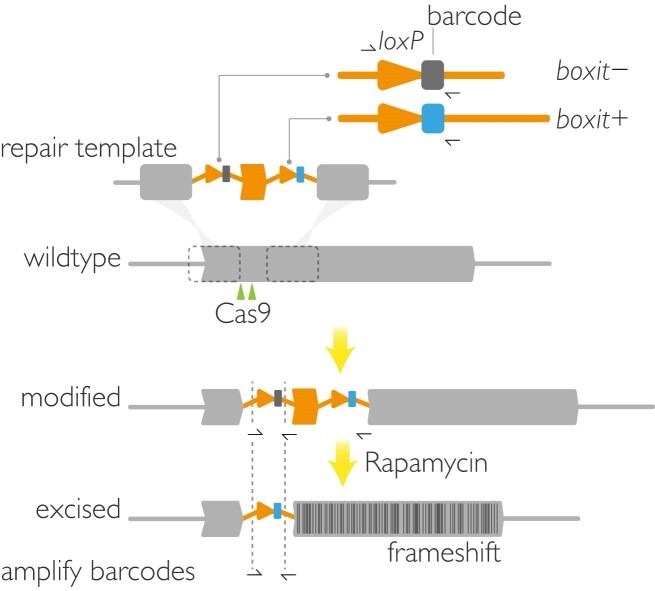
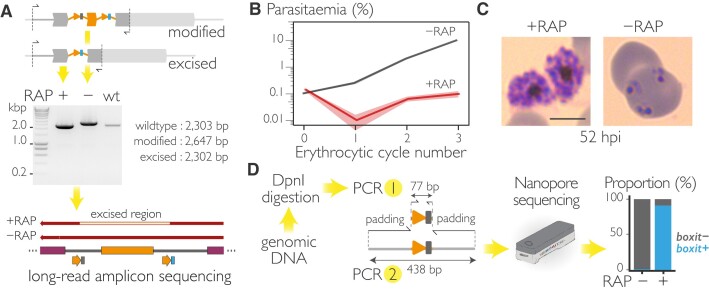
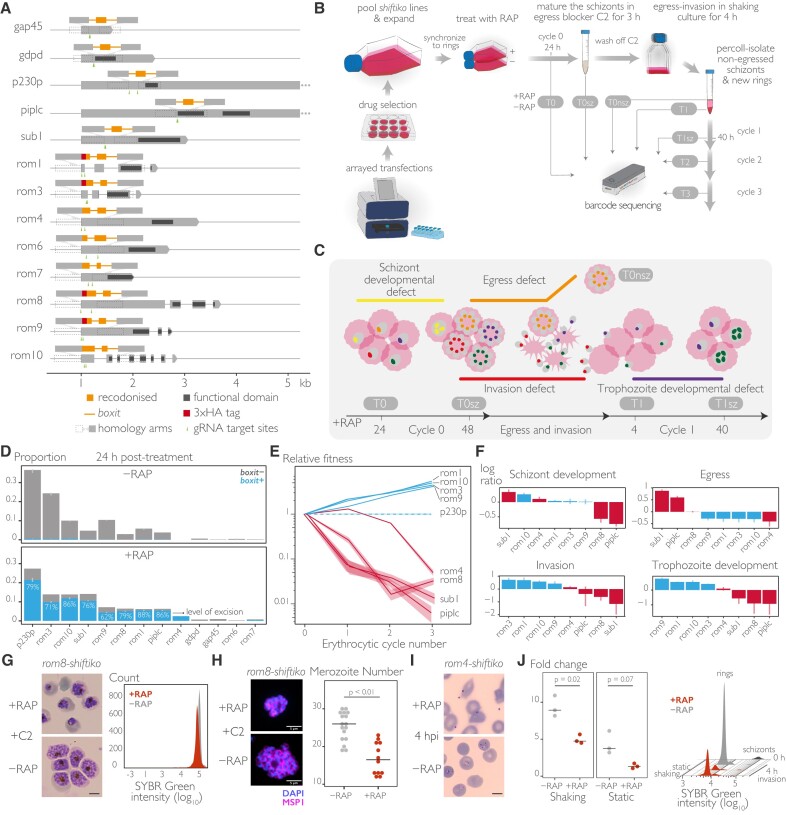
The malaria parasite needs nearly half of its genes to propagate normally within red blood cells. Inducible ways to interfere with gene expression like the DiCre-lox system are necessary to study the function of these essential genes. However, existing DiCre-lox strategies are not well-suited to be deployed at scale to study several genes simultaneously. To overcome this, we have developed SHIFTiKO (frameshift-based trackable inducible knockout), a novel scaleable strategy that uses short, easy-to-construct, barcoded repair templates to insert loxP sites around short regions in target genes. Induced DiCre-mediated excision of the flanked region causes a frameshift mutation resulting in genetic ablation of gene function. Dual DNA barcodes inserted into each mutant enables verification of successful modification and induced excision at each locus and collective phenotyping of the mutants, not only across multiple replication cycles to assess growth fitness but also within a single cycle to identify specific phenotypic impairments. As a proof of concept, we have applied SHIFTiKO to screen the functions of malarial rhomboid proteases, successfully identifying their blood stage-specific essentiality. SHIFTiKO thus offers a powerful platform to conduct inducible phenotypic screens to study essential gene function at scale in the malaria parasite.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- de Koning-Ward T.F., Gilson P.R., Crabb B.S.. Advances in molecular genetic systems in malaria. Nat. Rev. Microbiol. 2015; 13:373–387. - PubMed
-
- World Health Organization World Malaria Report 2023. 2023; Geneva: World Health Organization.
-
- Ghorbal M., Gorman M., Macpherson C.R., Martins R.M., Scherf A., Lopez-Rubio J.-J.. Genome editing in the human malaria parasite Plasmodium falciparum using the CRISPR-Cas9 system. Nat. Biotechnol. 2014; 32:819–821. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
