

Structural diversity of axonemes across mammalian motile cilia
- PMID: 39743588
- PMCID: PMC11779644
- DOI: 10.1038/s41586-024-08337-5
Structural diversity of axonemes across mammalian motile cilia
Abstract
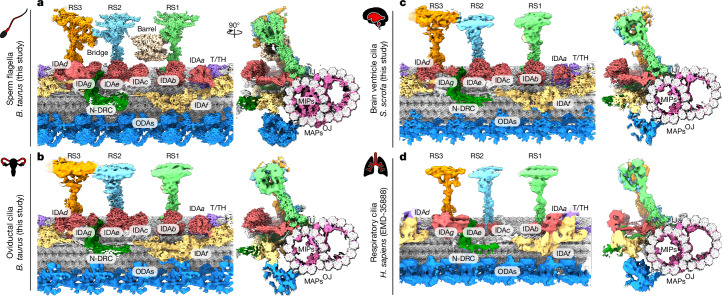
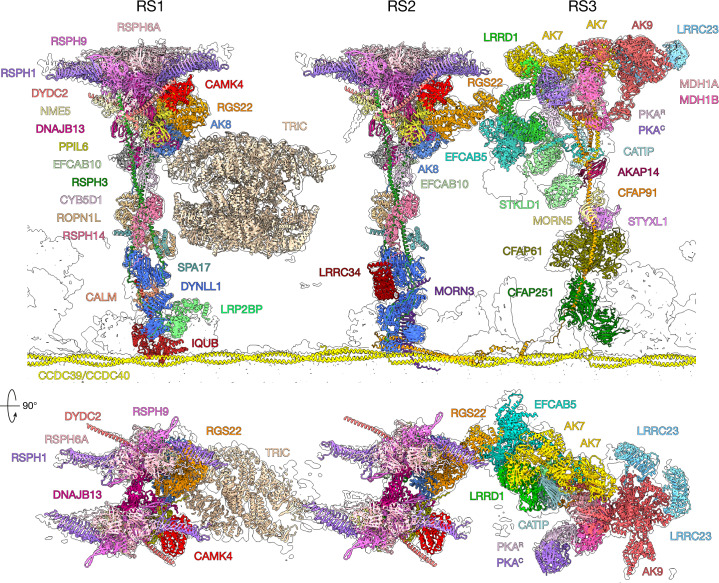
Reproduction, development and homeostasis depend on motile cilia, whose rhythmic beating is powered by a microtubule-based molecular machine called the axoneme. Although an atomic model of the axoneme is available for the alga Chlamydomonas reinhardtii1, structures of mammalian axonemes are incomplete1-5. Furthermore, we do not fully understand how molecular structures of axonemes vary across motile-ciliated cell types in the body. Here we use cryoelectron microscopy, cryoelectron tomography and proteomics to resolve the 96-nm modular repeat of axonemal doublet microtubules (DMTs) from both sperm flagella and epithelial cilia of the oviduct, brain ventricles and respiratory tract. We find that sperm DMTs are the most specialized, with epithelial cilia having only minor differences across tissues. We build a model of the mammalian sperm DMT, defining the positions and interactions of 181 proteins including 34 newly identified proteins. We elucidate the composition of radial spoke 3 and uncover binding sites of kinases associated with regeneration of ATP and regulation of ciliary motility. We discover a sperm-specific, axoneme-tethered T-complex protein ring complex (TRiC) chaperone that may contribute to construction or maintenance of the long flagella of mammalian sperm. We resolve axonemal dyneins in their prestroke states, illuminating conformational changes that occur during ciliary movement. Our results illustrate how elements of chemical and mechanical regulation are embedded within the axoneme, providing valuable resources for understanding the aetiology of ciliopathy and infertility, and exemplifying the discovery power of modern structural biology.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures













References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
