

Are we serologically prepared against an avian influenza pandemic and could seasonal flu vaccines help us?
- PMID: 39745389
- PMCID: PMC11796349
- DOI: 10.1128/mbio.03721-24
Are we serologically prepared against an avian influenza pandemic and could seasonal flu vaccines help us?
Abstract
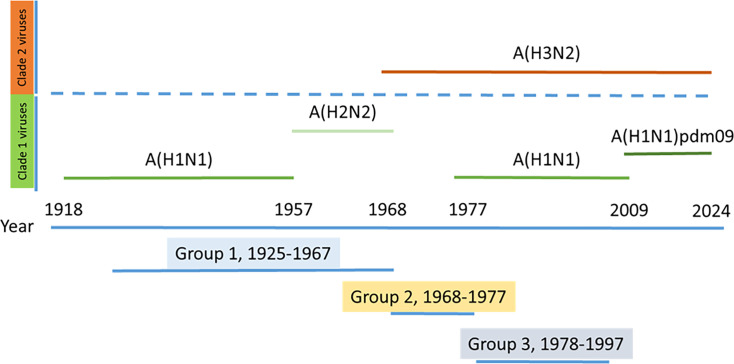
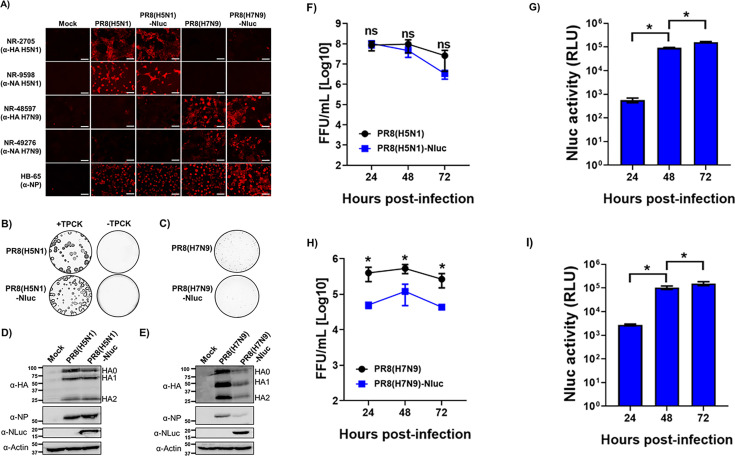
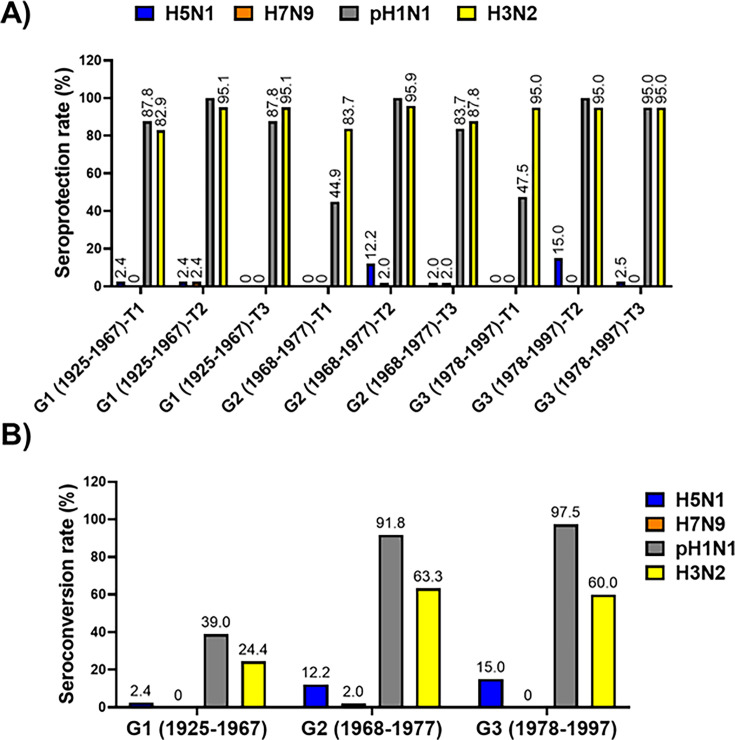
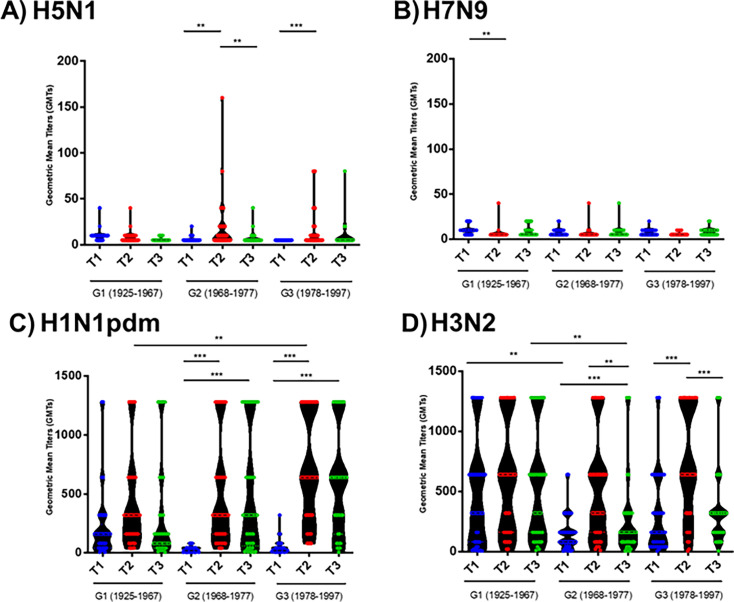
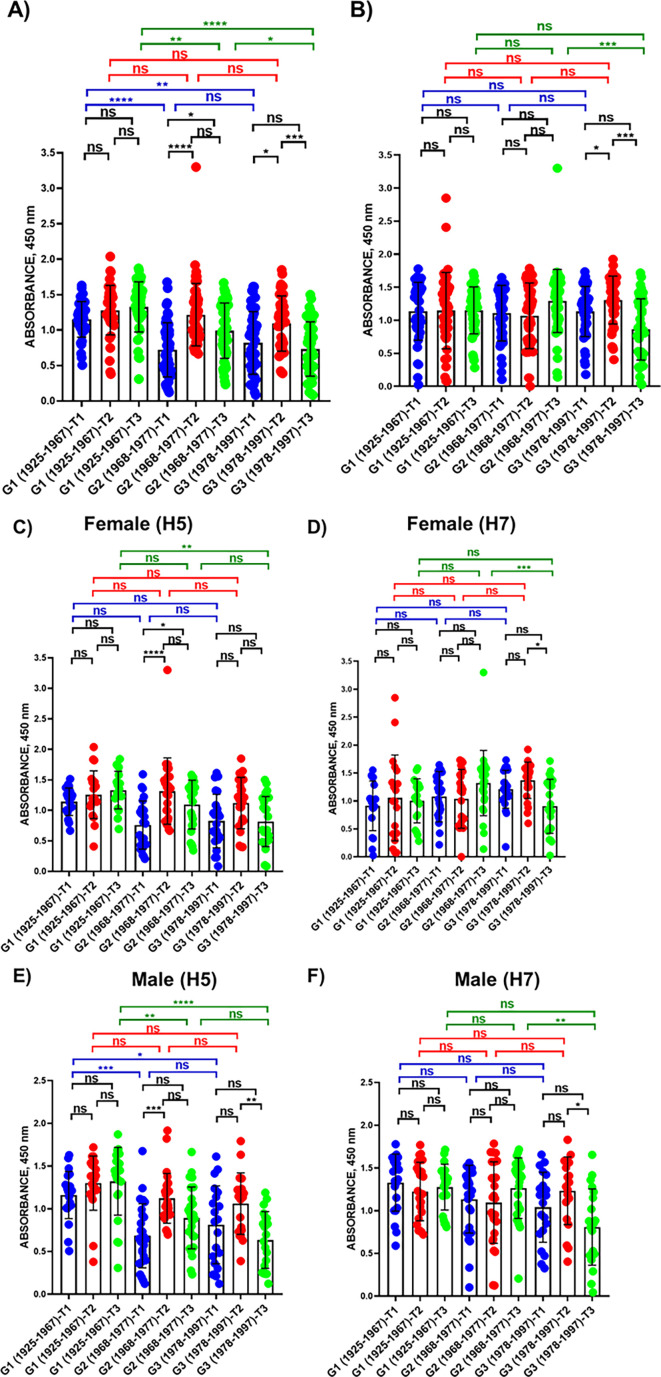
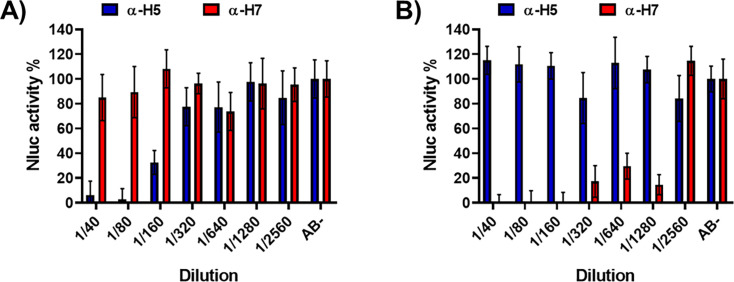
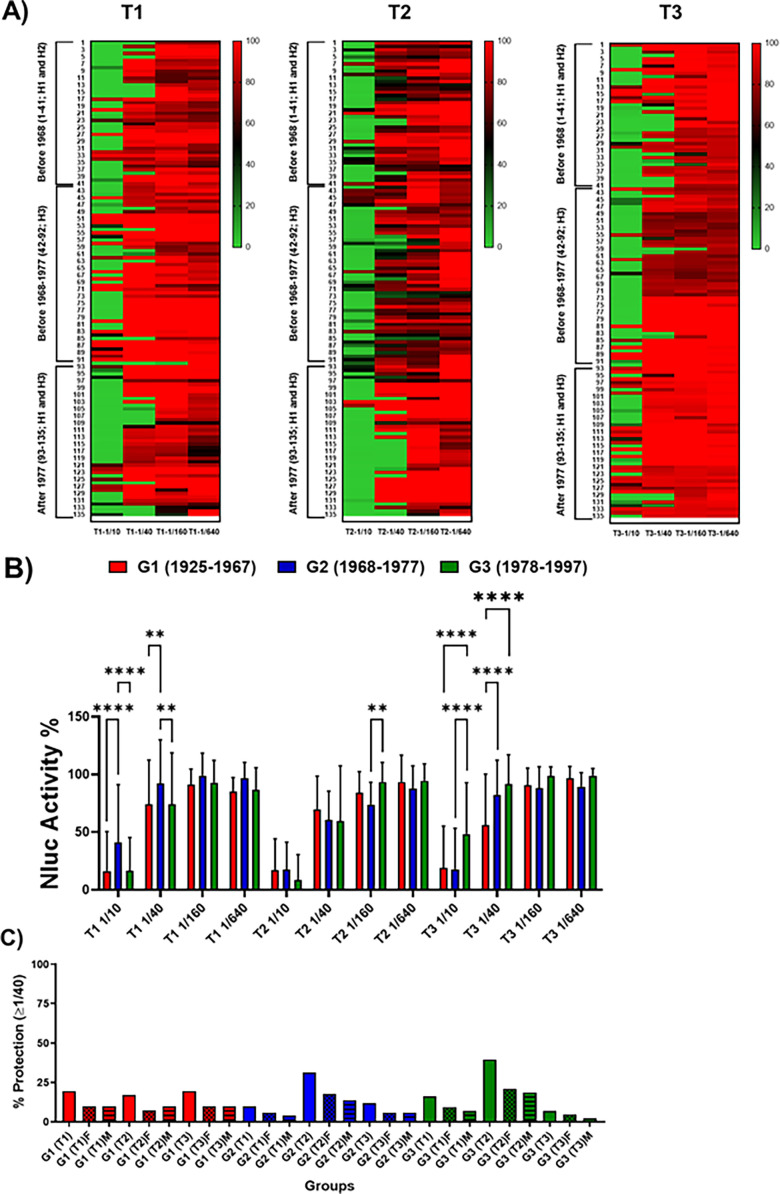
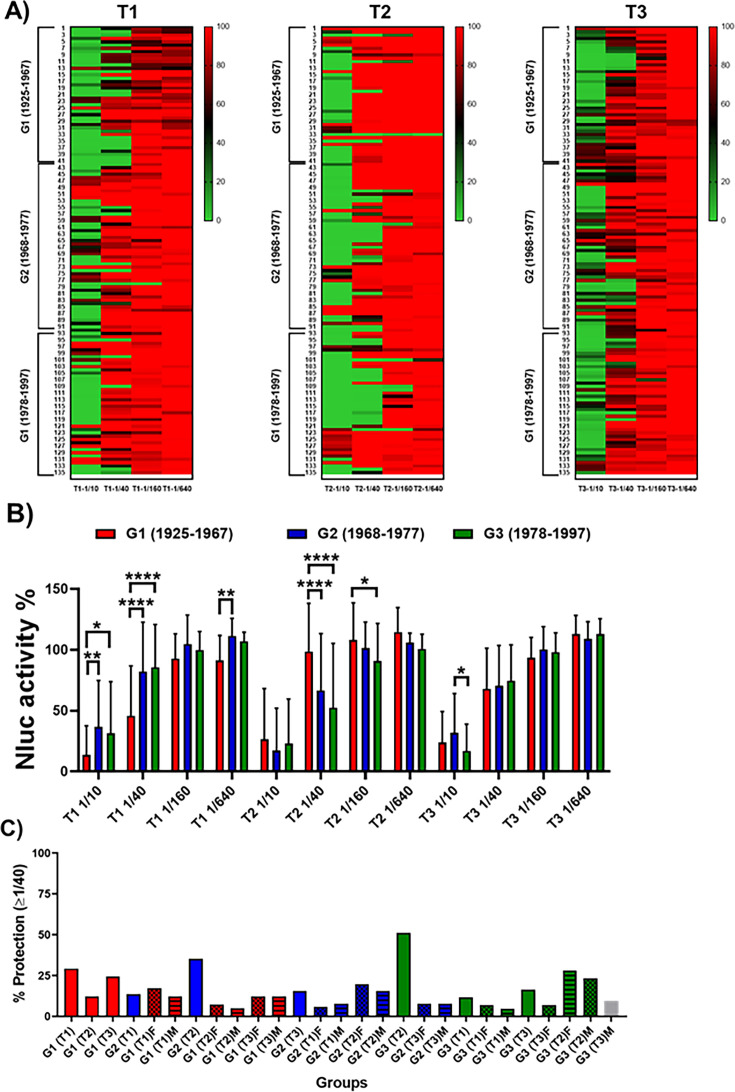
The current situation with H5N1 highly pathogenic avian influenza virus (HPAI) is causing a worldwide concern due to multiple outbreaks in wild birds, poultry, and mammals. Moreover, multiple zoonotic infections in humans have been reported. Importantly, HPAI H5N1 viruses with genetic markers of adaptation to mammals have been detected. Together with HPAI H5N1, avian influenza viruses H7N9 (high and low pathogenic) stand out due to their high mortality rates in humans. This raises the question of how prepared we are serologically and whether seasonal vaccines are capable of inducing protective immunity against these influenza subtypes. An observational study was conducted in which sera from people born between years 1925-1967, 1968-1977, and 1978-1997 were collected before or after 28 days or 6 months post-vaccination with an inactivated seasonal influenza vaccine. Then, hemagglutination inhibition, viral neutralization, and immunoassays were performed to assess the basal protective immunity of the population as well as the ability of seasonal influenza vaccines to induce protective responses. Our results indicate that subtype-specific serological protection against H5N1 and H7N9 in the representative Spanish population evaluated was limited or nonexistent. However, seasonal vaccination was able to increase the antibody titers to protective levels in a moderate percentage of people, probably due to cross-reactive responses. These findings demonstrate the importance of vaccination and suggest that seasonal influenza vaccines could be used as a first line of defense against an eventual pandemic caused by avian influenza viruses, to be followed immediately by the use of more specific pandemic vaccines.IMPORTANCEInfluenza A viruses (IAV) can infect and replicate in multiple mammalian and avian species. Avian influenza virus (AIV) is a highly contagious viral disease that occurs primarily in poultry and wild water birds. Due to the lack of population immunity in humans and ongoing evolution of AIV, there is a continuing risk that new IAV could emerge and rapidly spread worldwide, causing a pandemic, if the ability to transmit efficiently among humans was gained. The aim of this study is to analyze the basal protection and presence of antibodies against IAV H5N1 and H7N9 subtypes in the population from different ages. Moreover, we have evaluated the humoral response after immunization with a seasonal influenza vaccine. This study is strategically important to evaluate the level of population immunity that is a major factor when assessing the impact that an emerging IAV strain would have, and the role of seasonal vaccines to mitigate the effects of a pandemic.
Keywords: avian influenza; influenza A virus; influenza vaccine; nanoluciferase; neutralizing antibodies; pandemic; seroconversion; seroprotection.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
