

Dengue and Zika virus NS4B proteins differ in topology and in determinants of ER membrane protein complex dependency
- PMID: 39745435
- PMCID: PMC11852961
- DOI: 10.1128/jvi.01443-24
Dengue and Zika virus NS4B proteins differ in topology and in determinants of ER membrane protein complex dependency
Abstract
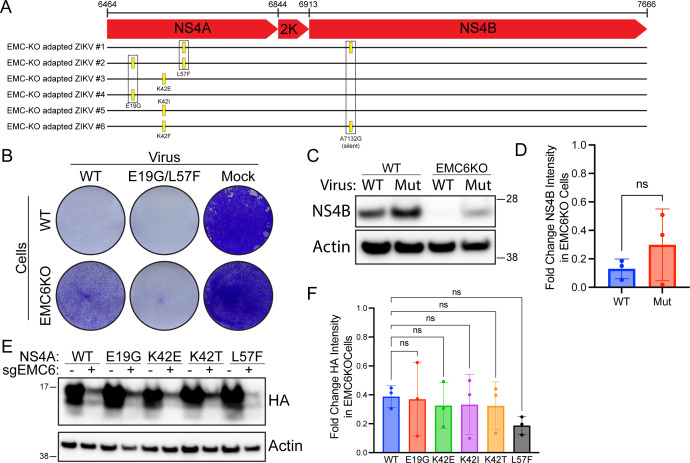
Flaviviruses utilize the cellular endoplasmic reticulum (ER) for all aspects of their lifecycle. Genome replication and other viral activities take place in structures called replication organelles (ROs), which are invaginations induced in the ER membrane. Among the required elements for RO formation is the biogenesis of viral nonstructural proteins NS4A and NS4B. We have previously shown that NS4A and NS4B from Dengue virus (DENV) and Zika virus (ZIKV) depend on the cellular ER membrane protein complex (EMC) for biogenesis. Here, we find that this dependency extends to the NS4A and NS4B proteins of Yellow Fever virus (YFV) and West Nile virus (WNV), which share similar computationally predicted membrane topologies. However, we demonstrate that ZIKV NS4B has different determinants of its dependency on the EMC than those for DENV NS4B, as well as a different membrane topology. Furthermore, we characterize mutant isolates of DENV and ZIKV that were serially passaged in EMC knockout cells and find that none are completely independent of the EMC for infection, and that mutant NS4B proteins remain sensitive to EMC depletion, suggesting a high genetic barrier to EMC depletion. Collectively, our findings are consistent with a model in which the EMC recognizes multiple determinants in the NS4B protein to support infection in several flaviviruses of critical public health importance.IMPORTANCEThe NS4A and NS4B proteins of flaviviruses are critically important to replication, but little is known about their function. It has been previously reported that the cellular EMC supports the biogenesis of NS4A and NS4B from Dengue and Zika virus. In this work, we demonstrate that this dependency on the EMC for NS4A and NS4B biogenesis extends to the West Nile and Yellow Fever viruses. Furthermore, we examine the features of ZIKV NS4B and find that its membrane topology of ZIKV NS4B and its determinants of dependency on the EMC are different from those previously described in DENV NS4B. Finally, we present evidence that there is a high genetic barrier for Dengue and Zika viruses to overcome EMC depletion.
Keywords: Dengue fever; Zika; flavivirus; virology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The ER Membrane Protein Complex Promotes Biogenesis of Dengue and Zika Virus Non-structural Multi-pass Transmembrane Proteins to Support Infection.Cell Rep. 2019 May 7;27(6):1666-1674.e4. doi: 10.1016/j.celrep.2019.04.051. Cell Rep. 2019. PMID: 31067454 Free PMC article.
-
Determinants in Nonstructural Protein 4A of Dengue Virus Required for RNA Replication and Replication Organelle Biogenesis.J Virol. 2021 Oct 13;95(21):e0131021. doi: 10.1128/JVI.01310-21. Epub 2021 Aug 11. J Virol. 2021. PMID: 34379504 Free PMC article.
-
Characterization of dengue virus NS4A and NS4B protein interaction.J Virol. 2015 Apr;89(7):3455-70. doi: 10.1128/JVI.03453-14. Epub 2015 Jan 7. J Virol. 2015. PMID: 25568208 Free PMC article.
-
Establishment and Application of Flavivirus Replicons.Adv Exp Med Biol. 2018;1062:165-173. doi: 10.1007/978-981-10-8727-1_12. Adv Exp Med Biol. 2018. PMID: 29845532 Review.
-
The Dengue Virus Replication Complex: From RNA Replication to Protein-Protein Interactions to Evasion of Innate Immunity.Adv Exp Med Biol. 2018;1062:115-129. doi: 10.1007/978-981-10-8727-1_9. Adv Exp Med Biol. 2018. PMID: 29845529 Review.
Cited by
-
Functional Roles and Host Interactions of Orthoflavivirus Non-Structural Proteins During Replication.Pathogens. 2025 Feb 12;14(2):184. doi: 10.3390/pathogens14020184. Pathogens. 2025. PMID: 40005559 Free PMC article. Review.
References
-
- Cortese M, Goellner S, Acosta EG, Neufeldt CJ, Oleksiuk O, Lampe M, Haselmann U, Funaya C, Schieber N, Ronchi P, Schorb M, Pruunsild P, Schwab Y, Chatel-Chaix L, Ruggieri A, Bartenschlager R. 2017. Ultrastructural characterization of Zika Virus replication factories. Cell Rep 18:2113–2123. doi:10.1016/j.celrep.2017.02.014 - DOI - PMC - PubMed
-
- Welsch S, Miller S, Romero-Brey I, Merz A, Bleck CKE, Walther P, Fuller SD, Antony C, Krijnse-Locker J, Bartenschlager R. 2009. Composition and three-dimensional architecture of the dengue virus replication and assembly sites. Cell Host Microbe 5:365–375. doi:10.1016/j.chom.2009.03.007 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- R21AI152865/HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- P30 DK034933/DK/NIDDK NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- R01 GM139823/GM/NIGMS NIH HHS/United States
- T32AI007528/HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
LinkOut - more resources
Full Text Sources
Medical
Research Materials
