

YBX1 is required for assembly of viral replication complexes of chikungunya virus and replication of multiple alphaviruses
- PMID: 39745458
- PMCID: PMC11852927
- DOI: 10.1128/jvi.02015-24
YBX1 is required for assembly of viral replication complexes of chikungunya virus and replication of multiple alphaviruses
Abstract
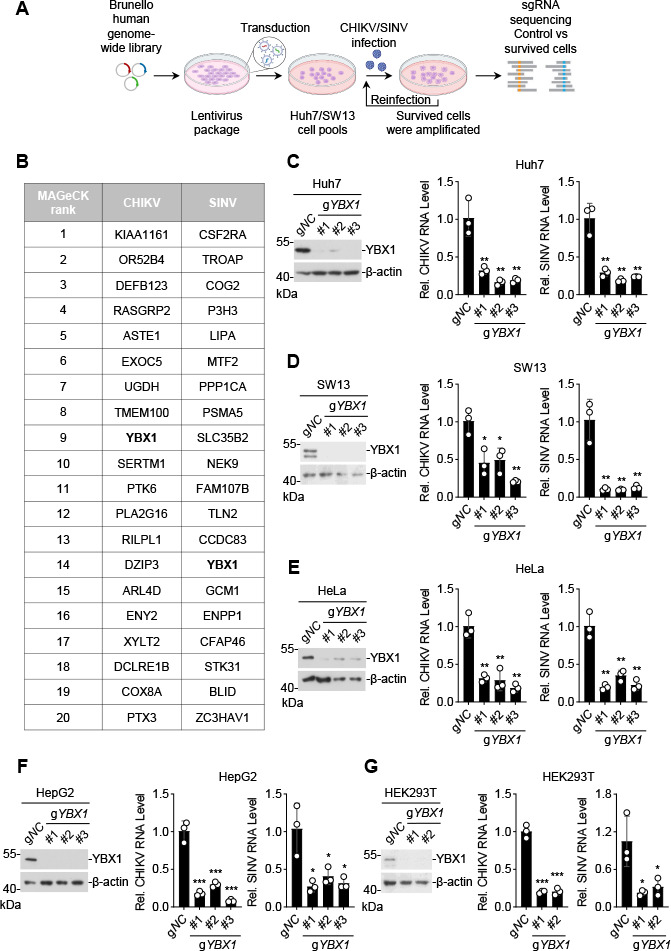
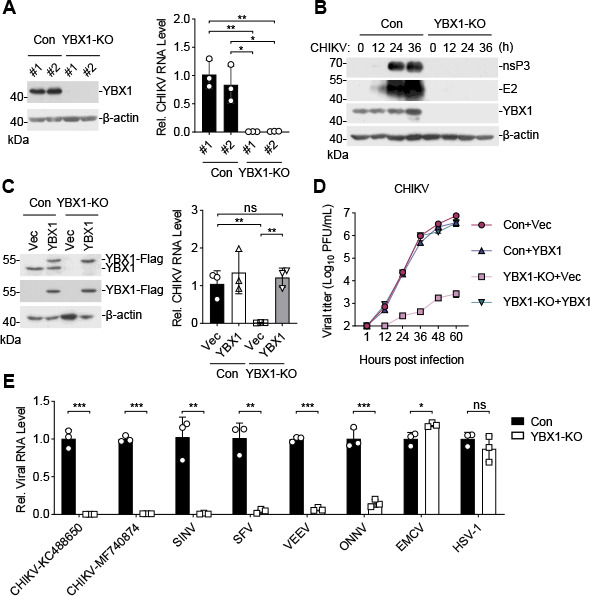
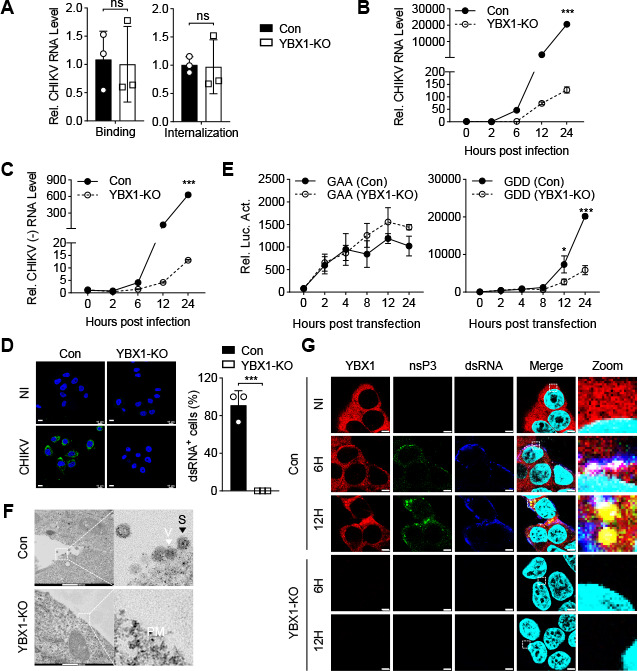
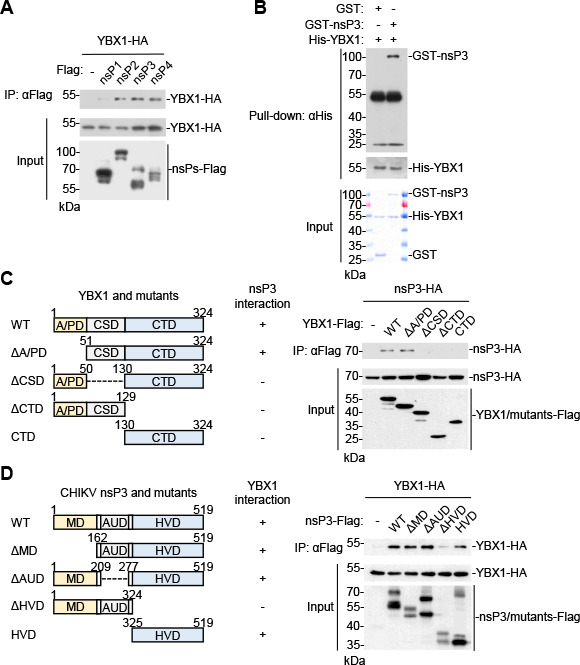
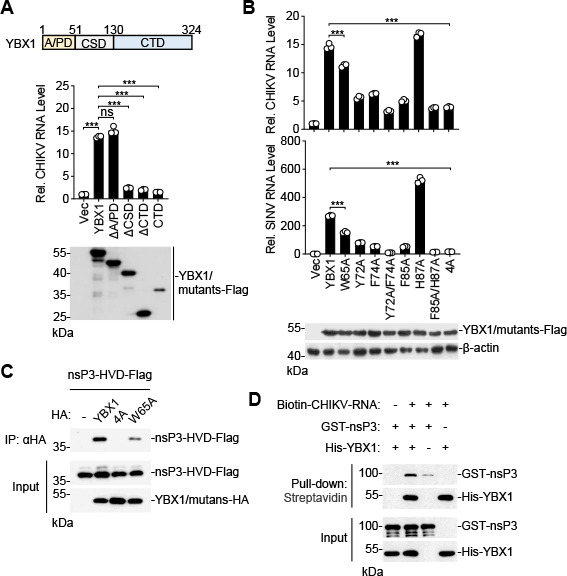
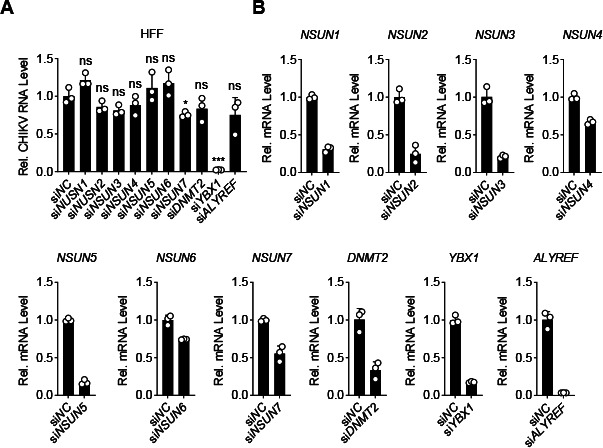
Chikungunya virus (CHIKV), an enveloped positive-sense RNA virus, is a member of the alphaviruses and cause fever and arthralgia in humans. We performed genome-wide CRISPR/Cas9-based screens and identified Y-box binding protein 1 (YBX1) as an essential cellular factor for CHIKV. Deficiency of YBX1 inhibited CHIKV RNA replication and impaired virus production. Upon CHIKV infection, YBX1 showed a striking re-localization to viral replication complexes (vRCs), where it co-localized with CHIKV nsP3 and dsRNA intermediates. YBX1 directly interacted with CHIKV nsP3, and mutation of the YBX1-binding motif in CHIKV nsP3 suppressed viral replication in host cells. Furthermore, YBX1 bound to viral RNA and increased the viral RNA-binding activity of CHIKV nsP3. Consistently, the RNA-binding activity of YBX1, as well as the ability of nsP3 to bind to YBX1, was required for efficient CHIKV replication. In addition to CHIKV, YBX1 was also essential for replication of all examined alphaviruses including the prototypic alphavirus. Our findings suggest that YBX1 acts as a scaffold for assembly of chikungunya vRCs and an important factor for replication of multiple alphaviruses, which may serve as a potential target for the development of anti-alphavirus therapies.IMPORTANCEAlphaviruses are a group of mosquito-transmitted, enveloped, positive-strand RNA viruses in the Togaviridae family. Most alphaviruses are important pathogens that continue to cause human disease ranging from severe and potentially fatal neurological disease to chronic arthritic disease on a global scale. Here, we found that YBX1 promotes binding of CHIKV genomic RNA to nsP3, which is a key component of the replication complex, and is therefore pivotal for CHIKV replication. Deficiency of YBX1 results in reduced replication of multiple alphaviruses, including arthritogenic and encephalitic alphaviruses. These findings suggest that YBX1 is an important cellular factor for multiple alphaviruses and a potential target for preventing alphavirus infections.
Keywords: YBX1; alphaviruses; cellular factor; viral replication.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- LaBeaud AD, Banda T, Brichard J, Muchiri EM, Mungai PL, Mutuku FM, Borland E, Gildengorin G, Pfeil S, Teng CY, Long K, Heise M, Powers AM, Kitron U, King CH. 2015. High rates of o'nyong nyong and Chikungunya virus transmission in coastal Kenya. PLoS Negl Trop Dis 9:e0003436. doi:10.1371/journal.pntd.0003436 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 2023YFC2305900/MOST | National Key Research and Development Program of China (NKPs)
- 2023YFC2306100/MOST | National Key Research and Development Program of China (NKPs)
- 32170930/MOST | National Natural Science Foundation of China (NSFC)
- U23A20168/MOST | National Natural Science Foundation of China (NSFC)
- 2022ACA005/Major Scientific and Technological Project of Hubei Province
LinkOut - more resources
Full Text Sources
Medical
Research Materials
