

Kir5.1 regulates Kir4.2 expression and is a key component of the 50-pS inwardly rectifying potassium channel in basolateral membrane of mouse proximal tubules
- PMID: 39745541
- PMCID: PMC12207947
- DOI: 10.1152/ajprenal.00178.2024
Kir5.1 regulates Kir4.2 expression and is a key component of the 50-pS inwardly rectifying potassium channel in basolateral membrane of mouse proximal tubules
Abstract
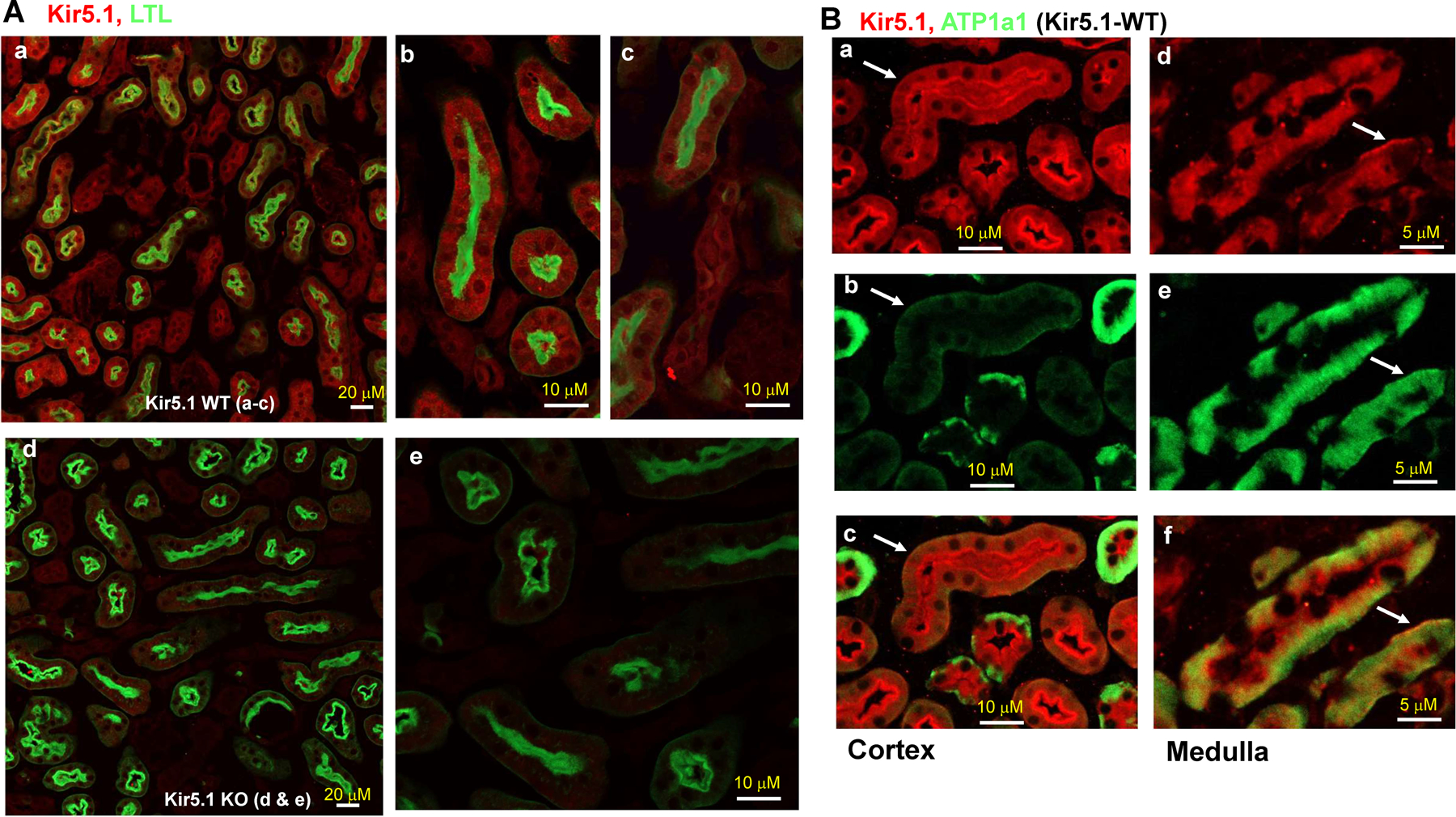
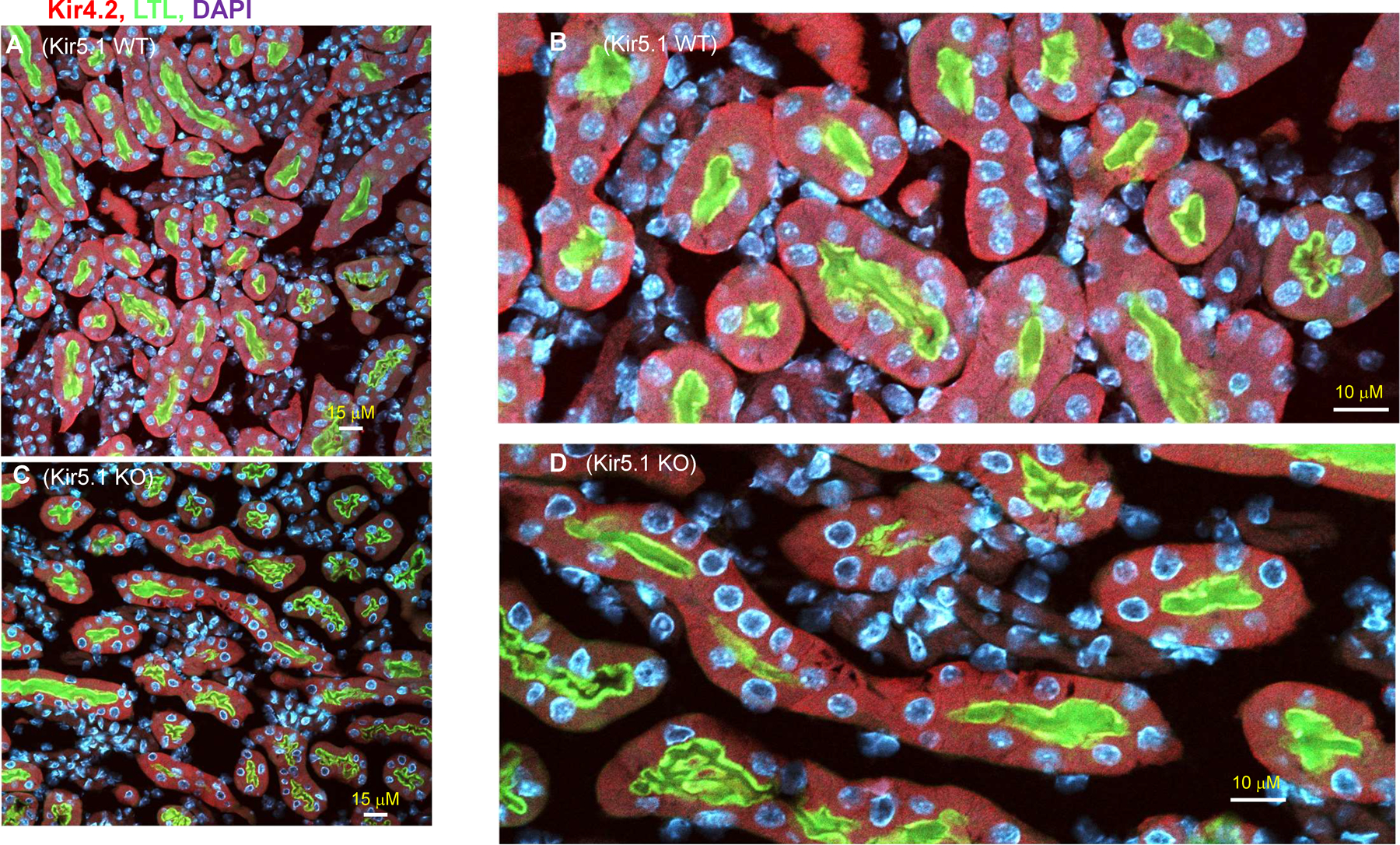
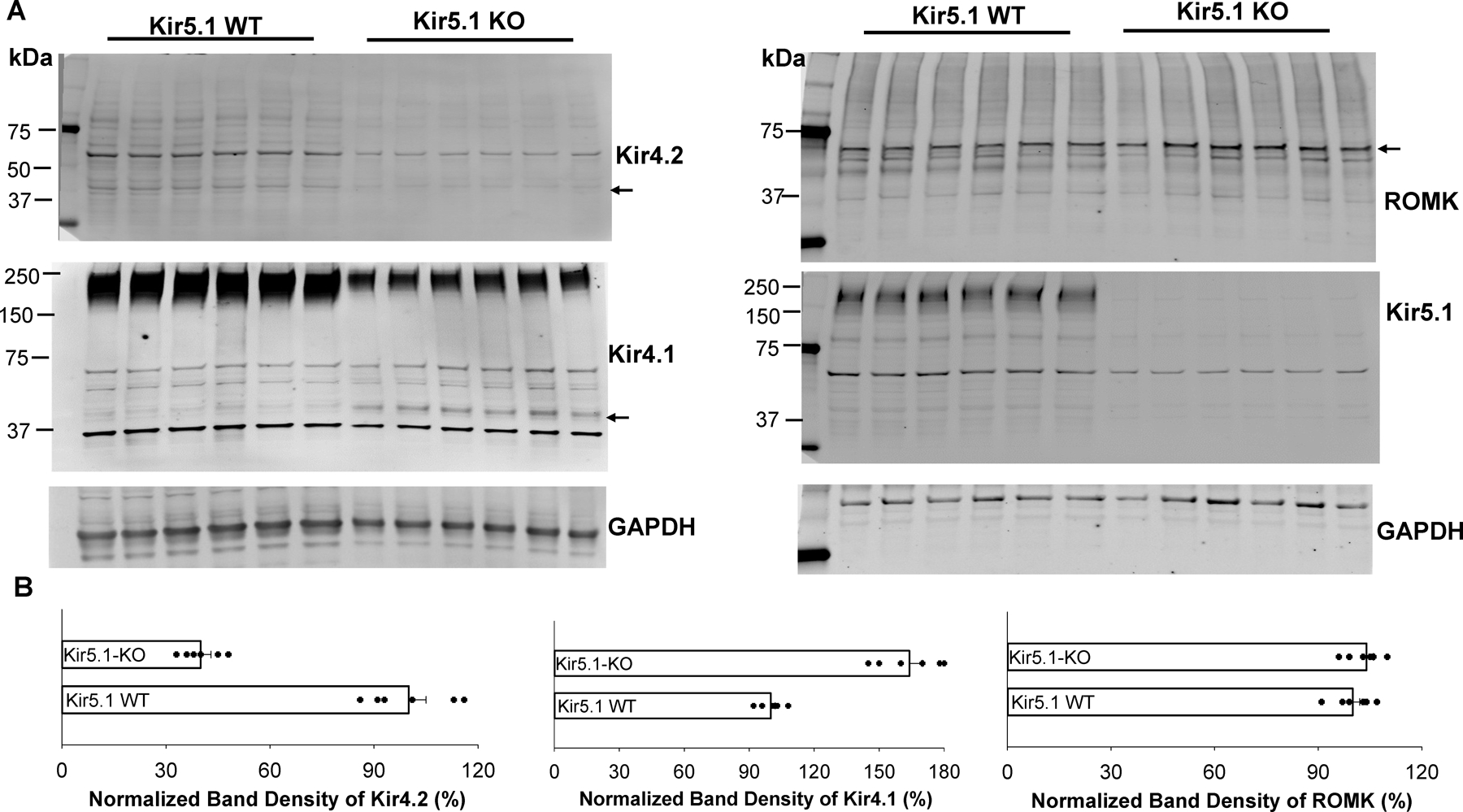
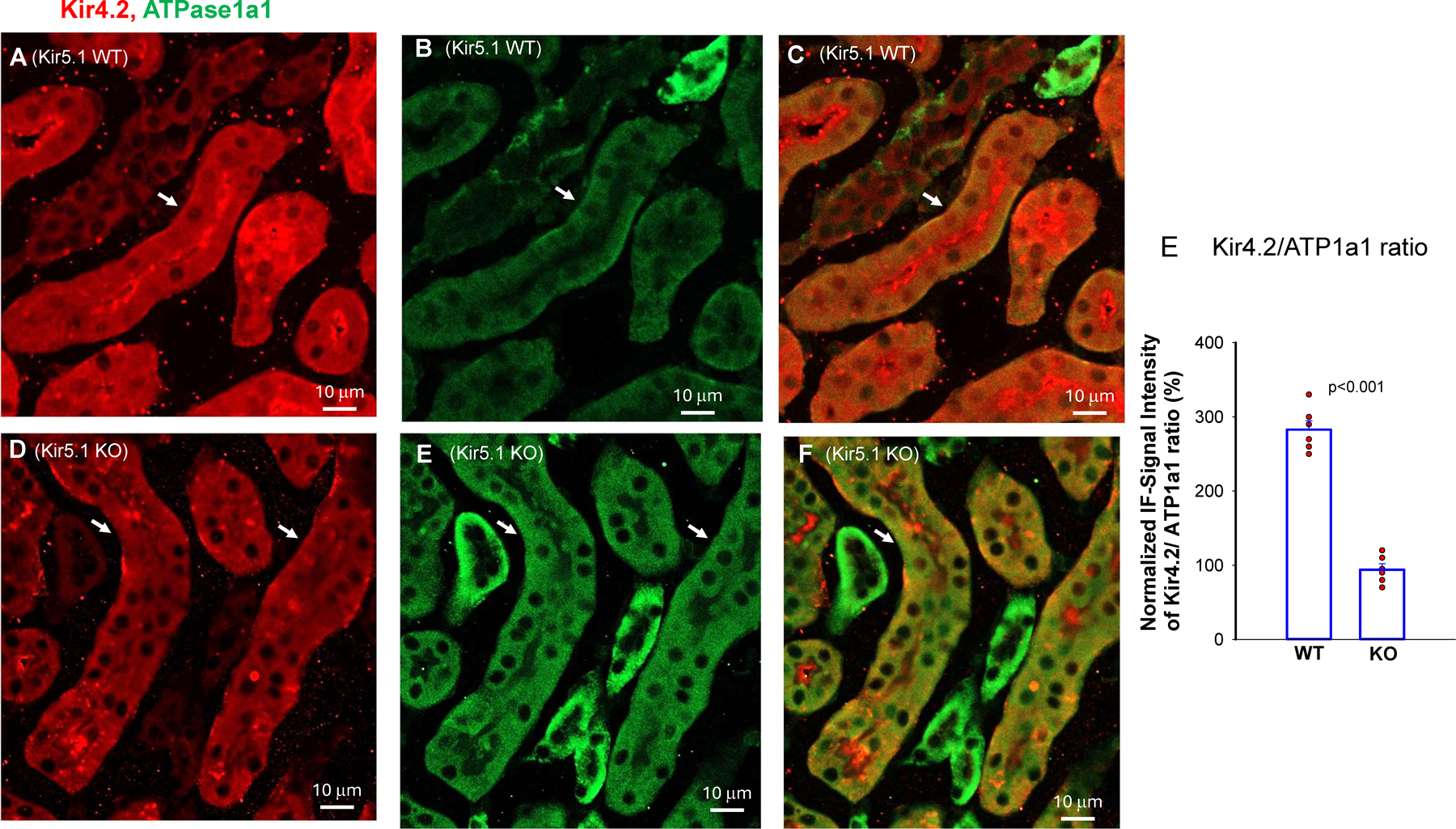
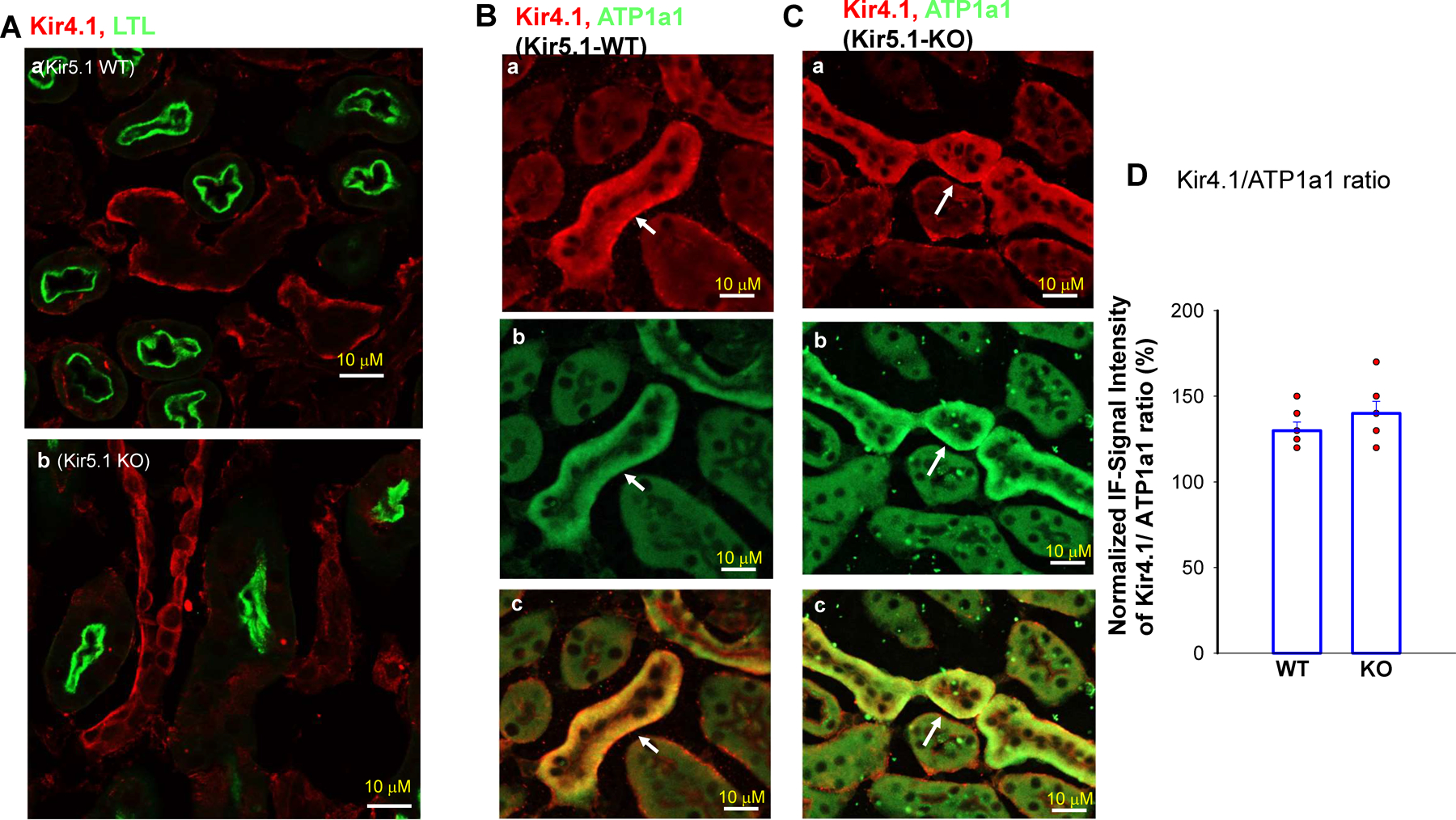
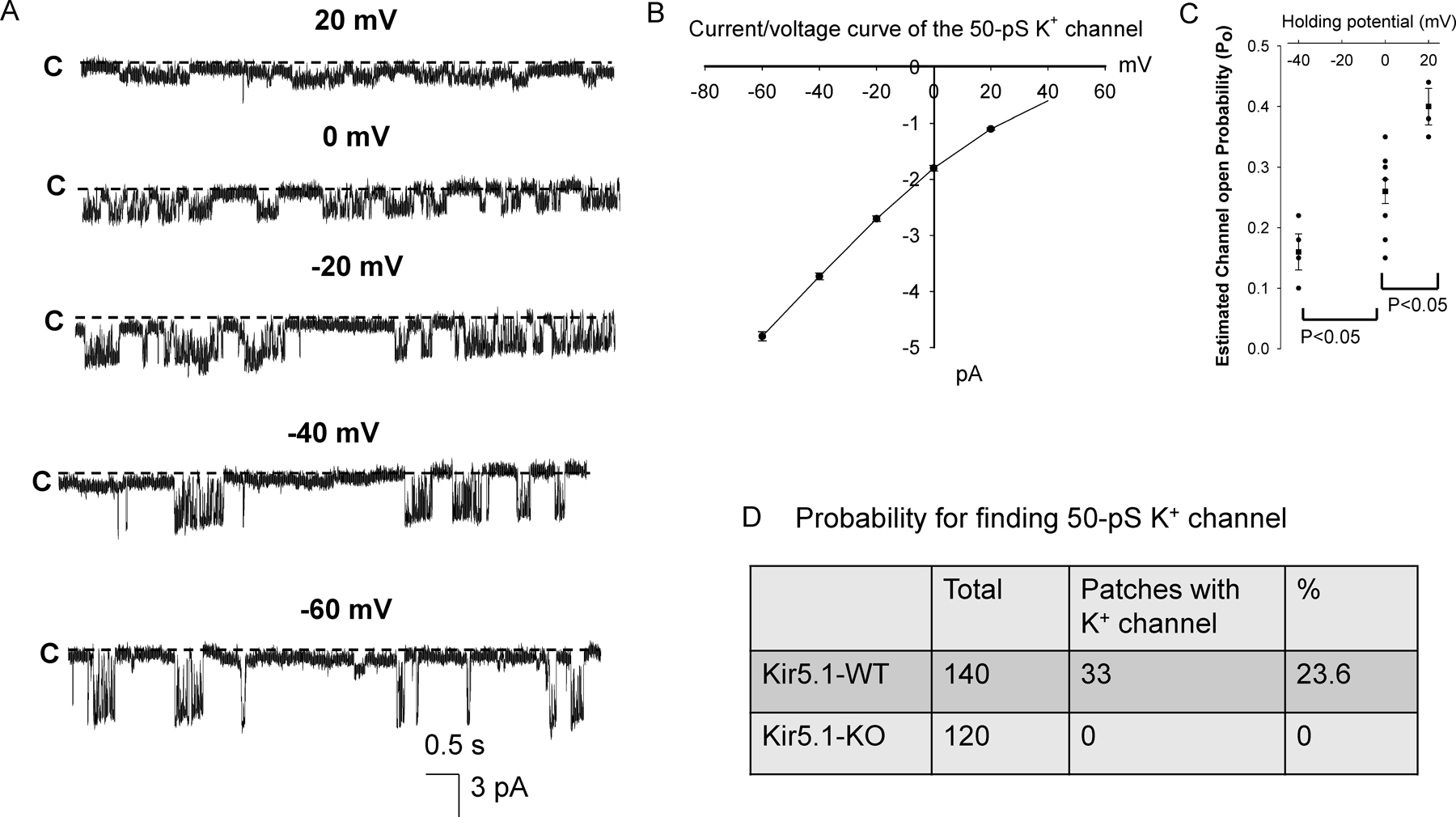
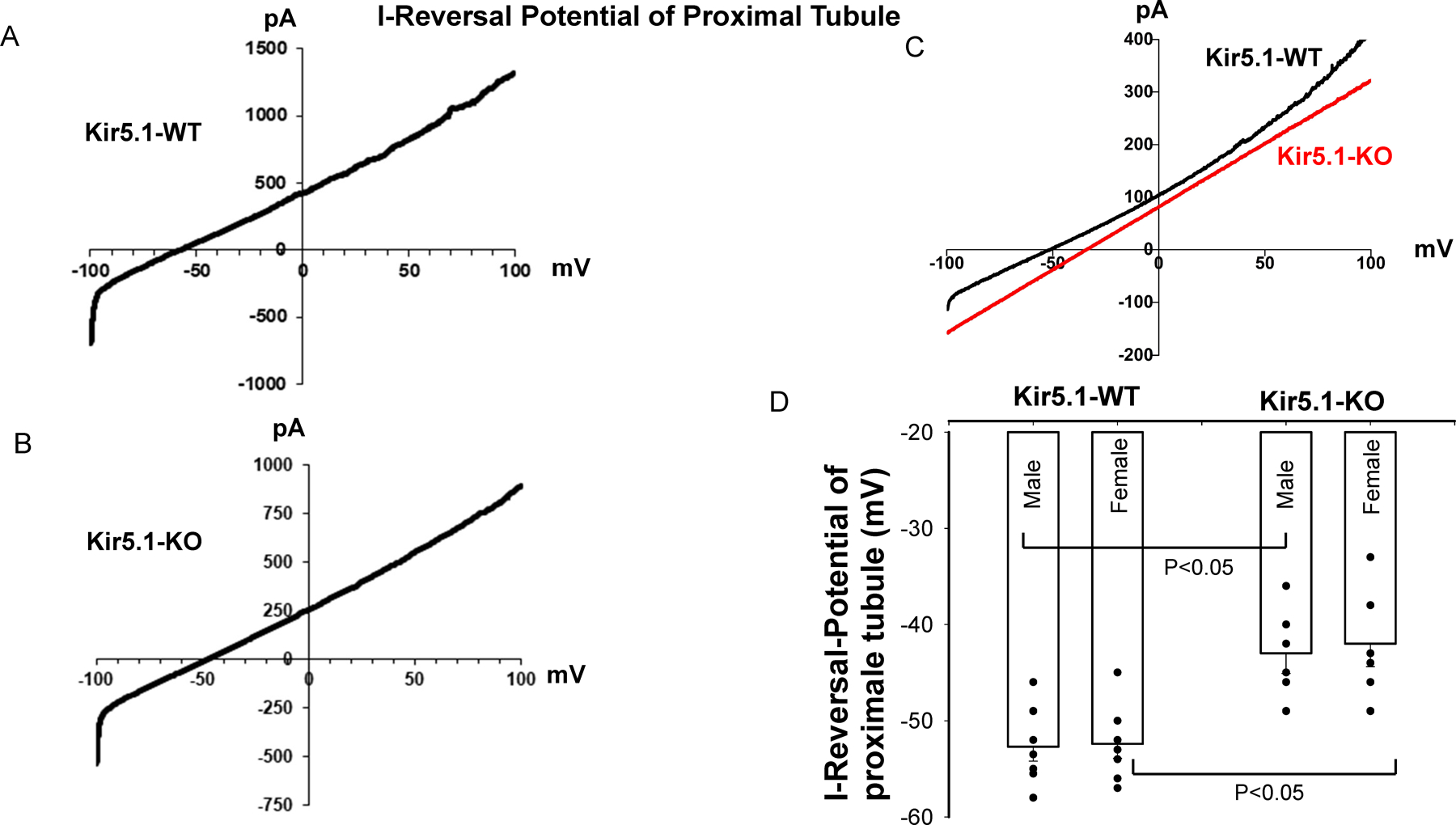
Kir5.1 encoded by Kcnj16 is an inwardly rectifying K+ channel subunit, and it possibly interacts with Kir4.2 subunit encoded by Kcnj15 for assembling a Kir4.2/Kir5.1 heterotetramer in the basolateral membrane of mouse proximal tubule. We now used patch clamp technique to examine basolateral K+ channels of mouse proximal tubule (PT) and an immunoblotting/immunofluorescence (IF) staining microscope to examine Kir4.2 expression in wild-type and Kir5.1-knockout mice. IF staining shows that Kir4.2 was exclusively expressed in the proximal tubule, whereas Kir5.1 was expressed in the proximal tubule and distal nephrons including distal convoluted tubule. Immunoblotting showed that the expression of Kir4.2 monomer was lower in Kir5.1-knockout mice than that in the wild-type mice. In contrast, Kir4.1 monomer expression was increased in Kir5.1 knockout mice. IF images further demonstrated that the basolateral membrane staining of Kir4.2 was significantly decreased in Kir5.1 knockout mice. This is in sharp contrast to Kir4.1, which also interacts with Kir5.1 in the distal nephron, and IF images show that Kir4.1 membrane expression was still visible and unchanged in Kir5.1 knockout mice. The single channel recording detected a 50-pS inwardly rectifying K+ channel, presumably a Kir4.2/Kir5.1 heterotetramer, in the basolateral membrane of the proximal tubule of Kir5.1 wild-type mice. However, this 50-pS K+ channel was completely absent in the basolateral membrane of the proximal tubule of Kir5.1 knockout mice. Moreover, the membrane potential of the proximal tubule was less negative in Kir5.1 knockout mice than wild-type mice. We conclude that Kir5.1 is essential for assembling basolateral 50-pS K+ channel in proximal tubule and that deletion of Kir5.1 decreased Kir4.2 expression in the proximal tubule thereby decreasing the basolateral K+ conductance and the membrane potentials.NEW & NOTEWORTHY Our study provides direct evidence for the notion that Kir5.1 is a key component of a 50-60 pS inwardly-rectifying-K+ channel, a main type K+ channel in the basolateral-membrane of PT. Also, we demonstrate that deletion of Kir5.1 decreased Kir4.2 protein expression including the basolateral-membrane in PT. Finally, depolarization of PT-membrane- potential in Kir5.1-knockout mice suggests that Kir4.2 alone is not able to sustain basolateral K+ conductance of the PT in the absence of Kir5.1.
Keywords: Kcnj10; Kcnj15; Kcnj16; potassium channel; proximal tubule.
Copyright © 2025 The Authors.
Conflict of interest statement
DISCLOSURES
None
Figures
Similar articles
-
Renal Tubule Nedd4-2 Deficiency Stimulates Kir4.1/Kir5.1 and Thiazide-Sensitive NaCl Cotransporter in Distal Convoluted Tubule.J Am Soc Nephrol. 2020 Jun;31(6):1226-1242. doi: 10.1681/ASN.2019090923. Epub 2020 Apr 15. J Am Soc Nephrol. 2020. PMID: 32295826 Free PMC article.
-
Deletion of renal Nedd4-2 abolishes the effect of high K+ intake on Kir4.1/Kir5.1 and NCC activity in the distal convoluted tubule.Am J Physiol Renal Physiol. 2021 Jul 1;321(1):F1-F11. doi: 10.1152/ajprenal.00072.2021. Epub 2021 May 24. Am J Physiol Renal Physiol. 2021. PMID: 34029145 Free PMC article.
-
Parkinson's disease-linked Kir4.2 mutation R28C leads to loss of ion channel function.J Physiol. 2025 Jun;603(12):3499-3518. doi: 10.1113/JP287046. Epub 2025 Jun 25. J Physiol. 2025. PMID: 40566643 Free PMC article.
-
Inwardly rectifying K+ channels 4.1 and 5.1 (Kir4.1/Kir5.1) in the renal distal nephron.Am J Physiol Cell Physiol. 2022 Aug 1;323(2):C277-C288. doi: 10.1152/ajpcell.00096.2022. Epub 2022 Jun 27. Am J Physiol Cell Physiol. 2022. PMID: 35759440 Free PMC article. Review.
-
Role of inwardly rectifying K+ channel 5.1 (Kir5.1) in the regulation of renal membrane transport.Curr Opin Nephrol Hypertens. 2022 Sep 1;31(5):479-485. doi: 10.1097/MNH.0000000000000817. Epub 2022 Jul 11. Curr Opin Nephrol Hypertens. 2022. PMID: 35894283 Review.
References
-
- Aronson PS. Mechanisms of active H+ secretion in the proximal tubule. Am J Physiol (Renal Fluid Electrolyte Physiol ) 245: F647–F659, 1983. - PubMed
-
- Aronson PS. The renal proximal tubule: A model for diversity of anion exchangers and stilbene-sensitive anion transporters. Ann Rev Physiol 51: 419–441, 1989. - PubMed
-
- Aronson PS, and Sacktor B. The Na+ gradient-dependent transport of D-glucose in renal brush border membranes. J Biol Chem 250: 6032–6039, 1975. - PubMed
-
- Wang W, Hebert SC, and Giebisch G. Renal K+ channels: structure and function. Ann Rev Physiol 59: 413–436, 1997. - PubMed
-
- Schultz SG. Homocellular regulatory mechanisms in sodium-transporting epithelia: avoidance of extinction by 'flush-through'. Am J Physiol (Renal Fluid Electrolyte Physiol ) 241: F579–F590, 1981. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous