

Strategic Acyl Carrier Protein Engineering Enables Functional Type II Polyketide Synthase Reconstitution In Vitro
- PMID: 39745931
- PMCID: PMC11744666
- DOI: 10.1021/acschembio.4c00678
Strategic Acyl Carrier Protein Engineering Enables Functional Type II Polyketide Synthase Reconstitution In Vitro
Abstract
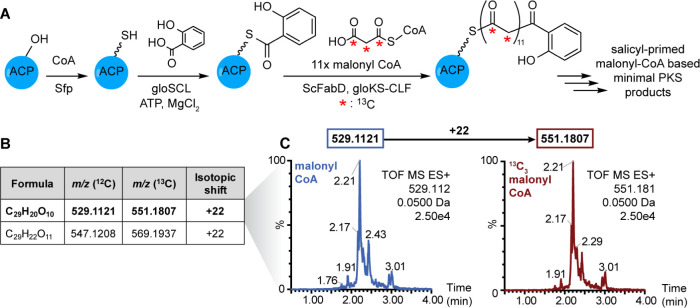
Microbial polyketides represent a structurally diverse class of secondary metabolites with medicinally relevant properties. Aromatic polyketides are produced by type II polyketide synthase (PKS) systems, each minimally composed of a ketosynthase-chain length factor (KS-CLF) and a phosphopantetheinylated acyl carrier protein (holo-ACP). Although type II PKSs are found throughout the bacterial kingdom, and despite their importance to strategic bioengineering, type II PKSs have not been well-studied in vitro. In cases where the KS-CLF can be accessed via E. coli heterologous expression, often the cognate ACPs are not activatable by the broad specificity Bacillus subtilis surfactin-producing phosphopantetheinyl transferase (PPTase) Sfp and, conversely, in systems where the ACP can be activated by Sfp, the corresponding KS-CLF is typically not readily obtained. Here, we report the high-yield heterologous expression of both cyanobacterial Gloeocapsa sp. PCC 7428 minimal type II PKS (gloPKS) components in E. coli, which allowed us to study this minimal type II PKS in vitro. Initially, neither the cognate PPTase nor Sfp converted gloACP to its active holo state. However, by examining sequence differences between Sfp-compatible and -incompatible ACPs, we identified two conserved residues in gloACP that, when mutated, enabled high-yield phosphopantetheinylation of gloACP by Sfp. Using analogous mutations, other previously Sfp-incompatible type II PKS ACPs from different bacterial phyla were also rendered activatable by Sfp. This demonstrates the generalizability of our approach and breaks down a longstanding barrier to type II PKS studies and the exploration of complex biosynthetic pathways.
Conflict of interest statement
The authors declare no competing financial interest.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
