

Structures of Saccharolobus solfataricus initiation complexes with leaderless mRNAs highlight archaeal features and eukaryotic proximity
- PMID: 39753558
- PMCID: PMC11698992
- DOI: 10.1038/s41467-024-55718-5
Structures of Saccharolobus solfataricus initiation complexes with leaderless mRNAs highlight archaeal features and eukaryotic proximity
Abstract
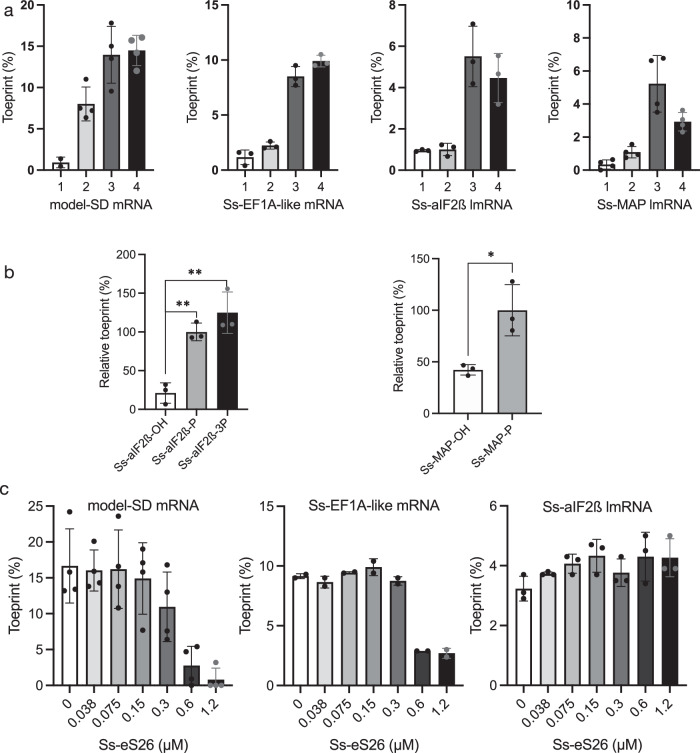
The archaeal ribosome is of the eukaryotic type. TACK and Asgard superphyla, the closest relatives of eukaryotes, have ribosomes containing eukaryotic ribosomal proteins not found in other archaea, eS25, eS26 and eS30. Here, we investigate the case of Saccharolobus solfataricus, a TACK crenarchaeon, using mainly leaderless mRNAs. We characterize the small ribosomal subunit of S. solfataricus bound to SD-leadered or leaderless mRNAs. Cryo-EM structures show eS25, eS26 and eS30 bound to the small subunit. We identify two ribosomal proteins, aS33 and aS34, and an additional domain of eS6. Leaderless mRNAs are bound to the small subunit with contribution of their 5'-triphosphate group. Archaeal eS26 binds to the mRNA exit channel wrapped around the 3' end of rRNA, as in eukaryotes. Its position is not compatible with an SD:antiSD duplex. Our results suggest a positive role of eS26 in leaderless mRNAs translation and possible evolutionary routes from archaeal to eukaryotic translation.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interest.
Figures







References
-
- Forterre, P., Brochier, C. & Philippe, H. Evolution of the Archaea. Theor. Popul Biol.61, 409–422 (2002). - PubMed
-
- Borrel, G., Brugère, J. F., Gribaldo, S., Schmitz, R. A. & Moissl-Eichinger, C. The host-associated archaeome. Nat. Rev. Microbiol18, 622–636 (2020). - PubMed
-
- Garcia, P. S., Gribaldo, S. & Borrel, G. Diversity and evolution of methane-related pathways in Archaea. Annu Rev. Microbiol76, 727–755 (2022). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
