

Single cell and spatial analysis of immune-hot and immune-cold tumours identifies fibroblast subtypes associated with distinct immunological niches and positive immunotherapy response
- PMID: 39757146
- PMCID: PMC11702232
- DOI: 10.1186/s12943-024-02191-9
Single cell and spatial analysis of immune-hot and immune-cold tumours identifies fibroblast subtypes associated with distinct immunological niches and positive immunotherapy response
Abstract
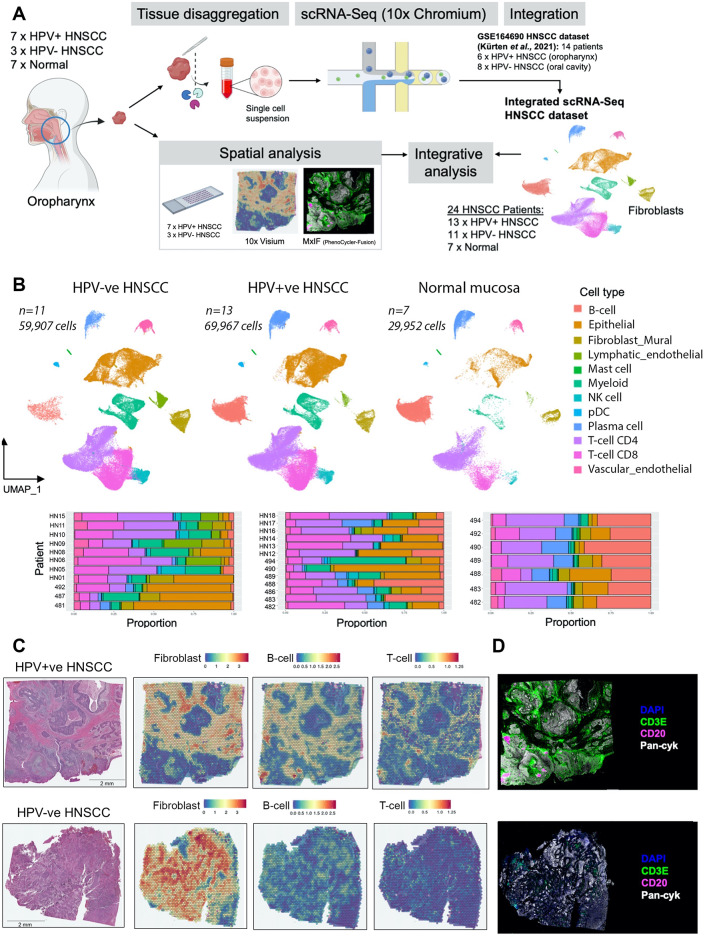
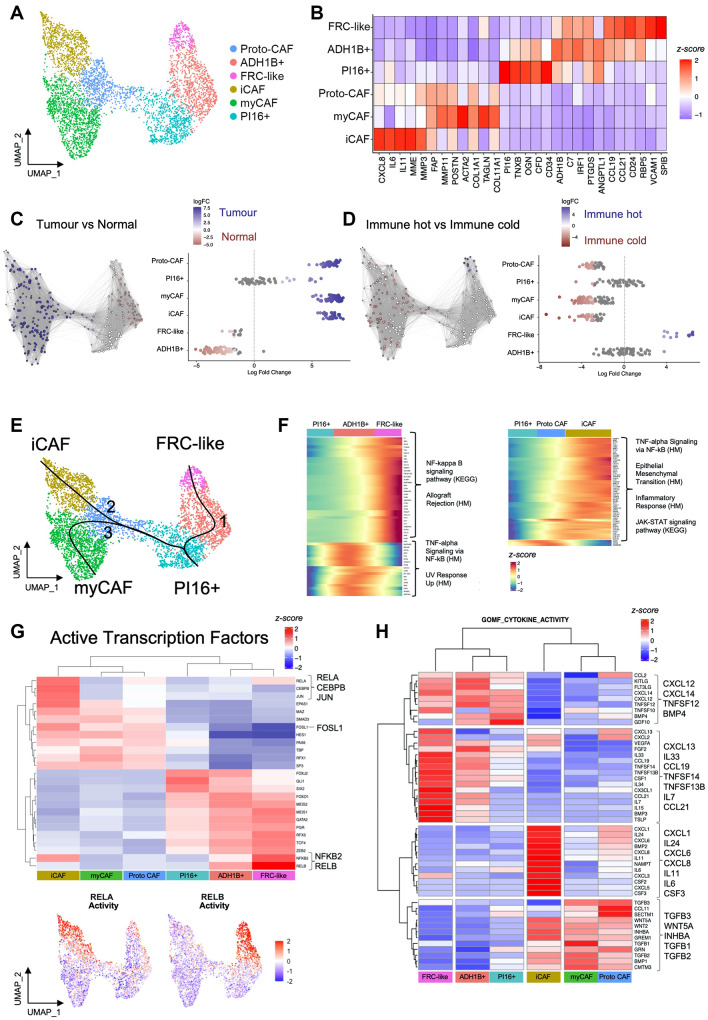
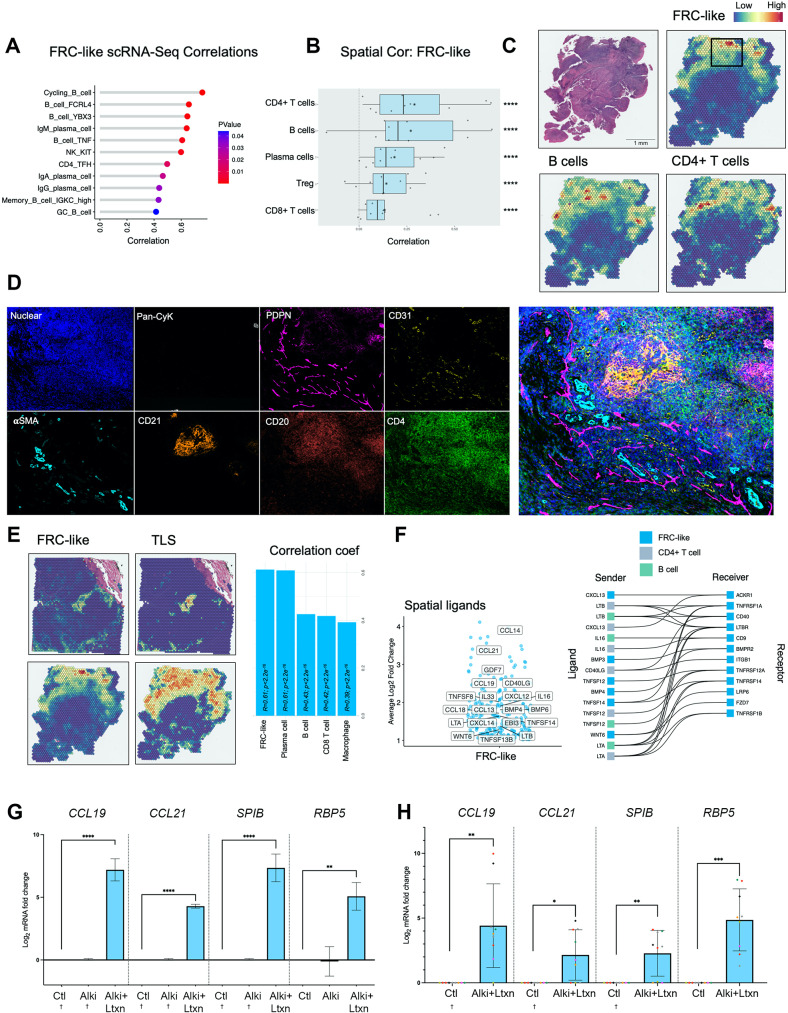
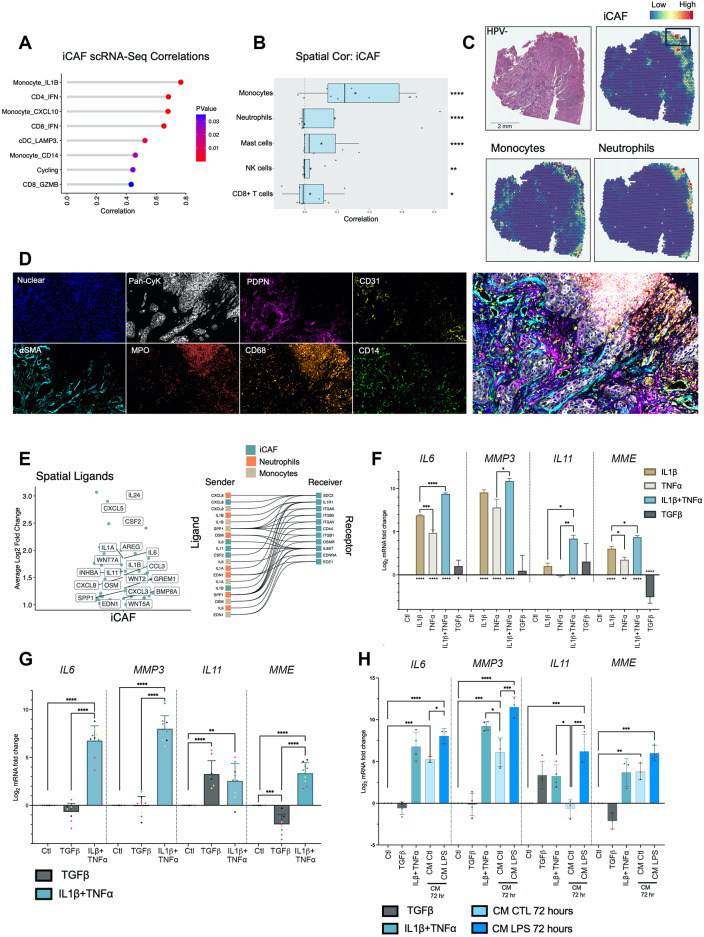
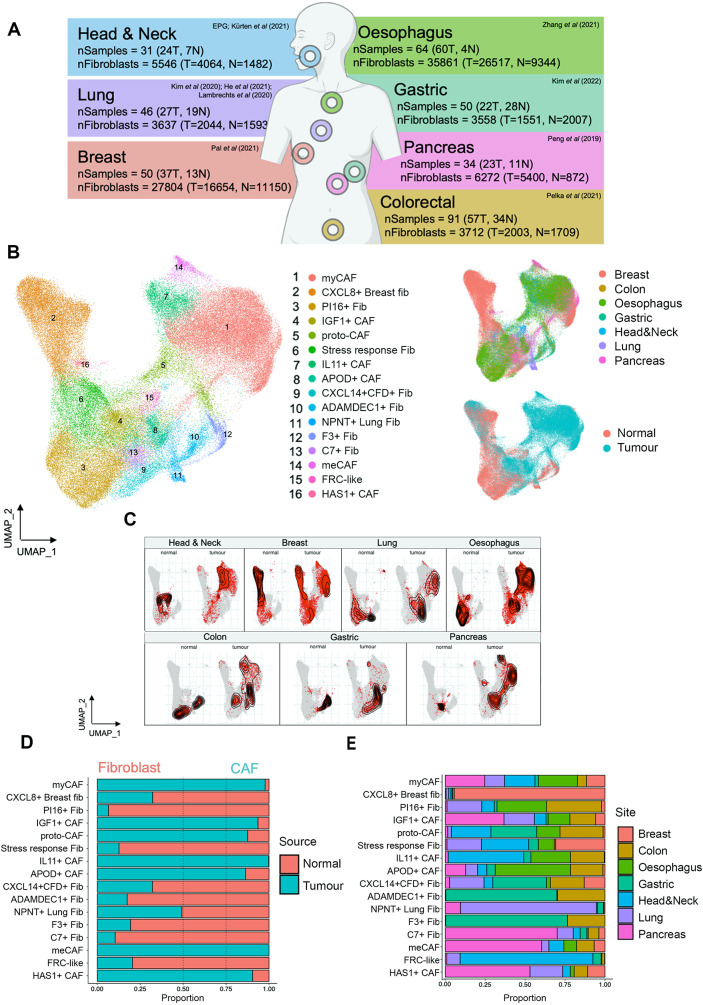
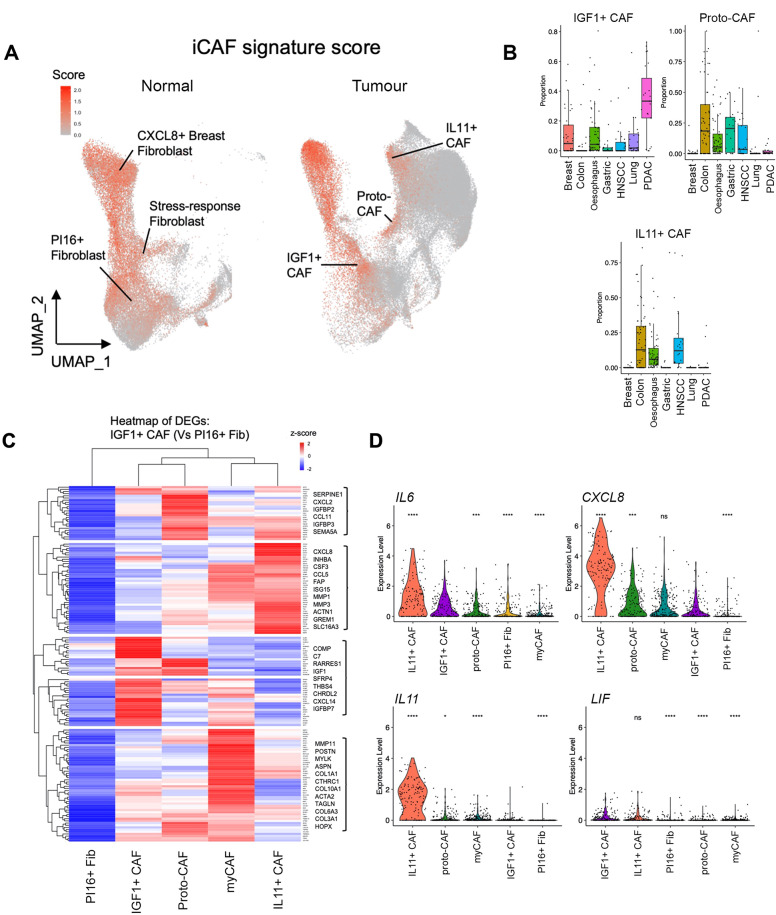
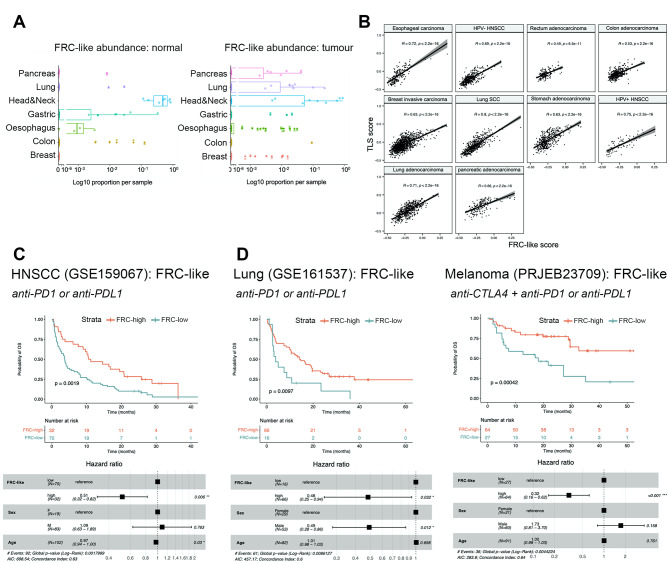
Cancer-associated Fibroblasts (CAFs) have emerged as critical regulators of anti-tumour immunity, with both beneficial and detrimental properties that remain poorly characterised. To investigate this, we performed single-cell and spatial transcriptomic analysis, comparing head & neck squamous cell carcinoma (HNSCC) subgroups, which although heterogenous, can be considered broadly immune-hot and immune-cold (human papillomavirus [HPV]+ve and HPV-ve tumours respectively). This identified six fibroblast subpopulations, including two with immunomodulatory gene expression profiles (IL-11 + inflammatory [i]CAF and CCL19 + fibroblastic reticular cell [FRC]-like). IL-11 + iCAF were spatially associated with inflammatory monocytes and regulated in vitro through synergistic activation of canonical NF-κB signalling by IL-1β and TNF-α. FRC-like were enriched in immune-hot HPV+ve tumours, associated with CD4 + T-cells and B-cells in tertiary lymphoid structures and regulated through non-canonical NF-κB signalling via lymphotoxin. Pan-cancer analysis revealed several 'iCAF' subgroups present in both normal and cancer tissues; IL11 + iCAF were found in cancers from the gastrointestinal (GI) tract and transcriptomically distinct from iCAFs previously described in pancreatic and breast cancers with greater inflammatory properties; FRC-like fibroblasts were present at low frequencies in all tumour types, and were associated with significantly better survival in patients receiving checkpoint immunotherapy. This work clarifies and expands current literature on immunomodulatory CAFs, highlighting links with important immunological niches.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethical approval: Ethical approval for the study was obtained through the UK National Research Ethics Service (South Central - Hampshire B Research Ethics Committee) and written informed consent was obtained from all subjects (REC No. 09/H0501/90). Competing interests: The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
