

Disrupting the transmembrane domain interface between PMP22 and MPZ causes peripheral neuropathy
- PMID: 39759075
- PMCID: PMC11700639
- DOI: 10.1016/j.isci.2024.110989
Disrupting the transmembrane domain interface between PMP22 and MPZ causes peripheral neuropathy
Abstract
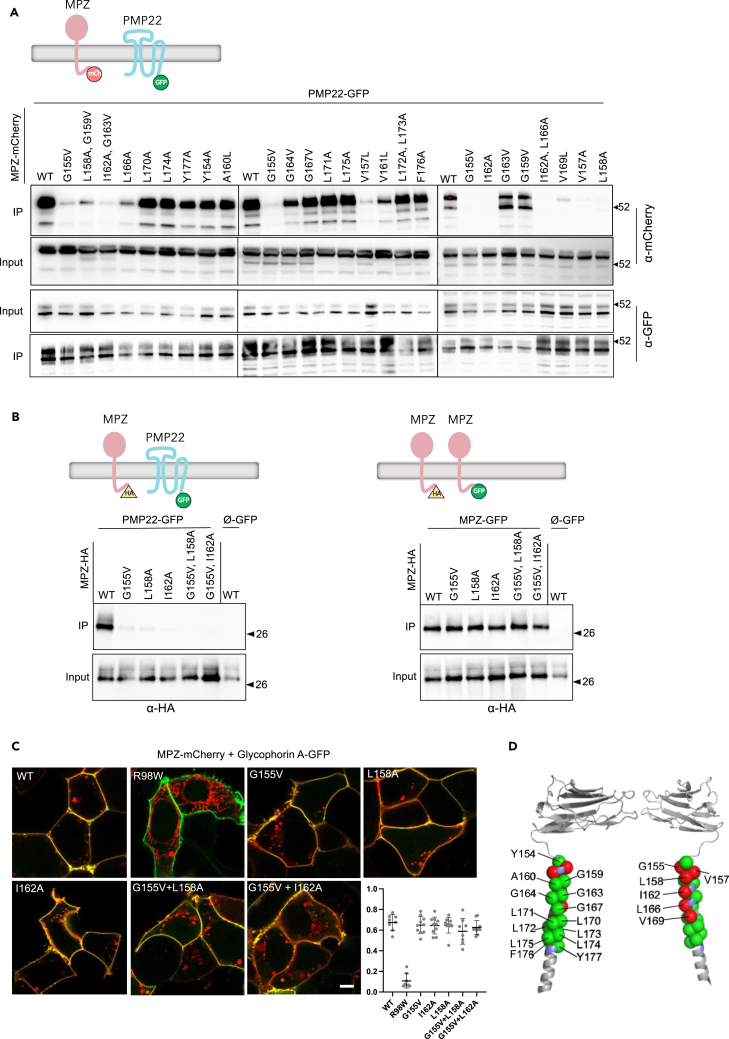
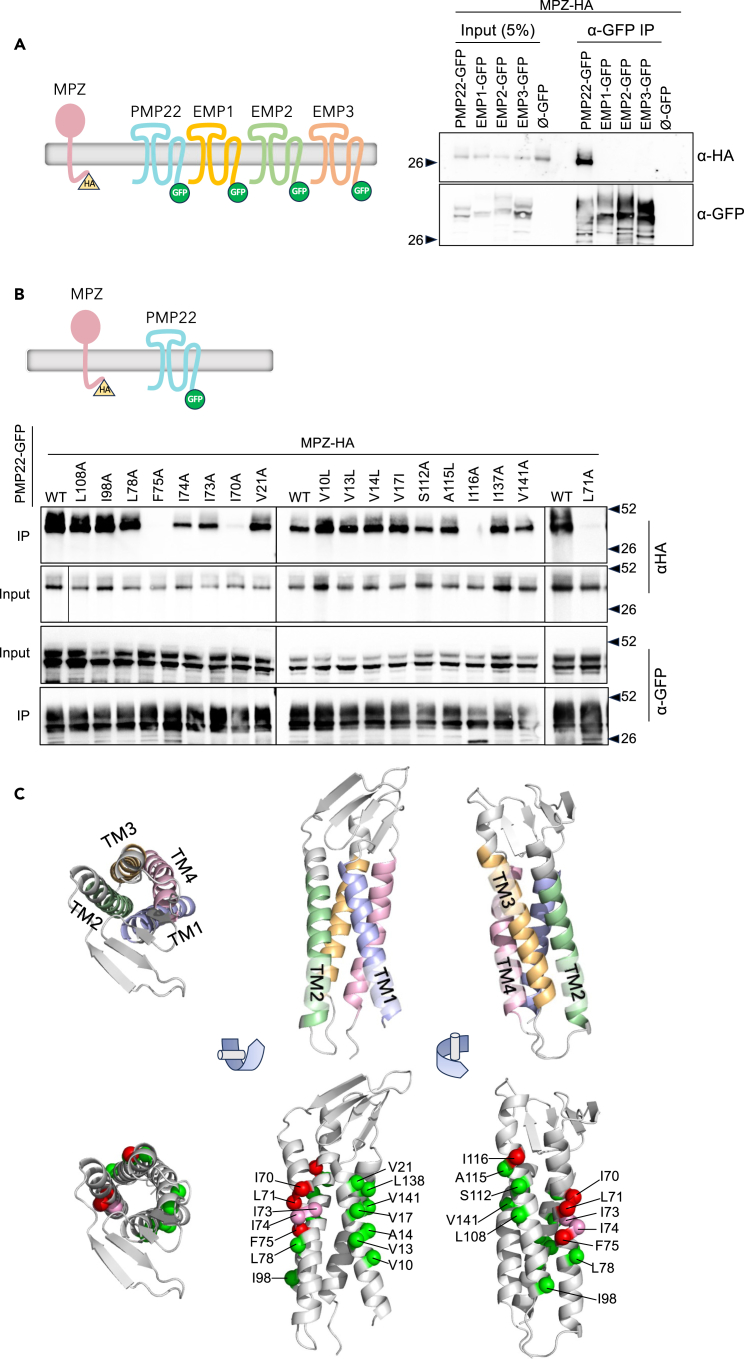
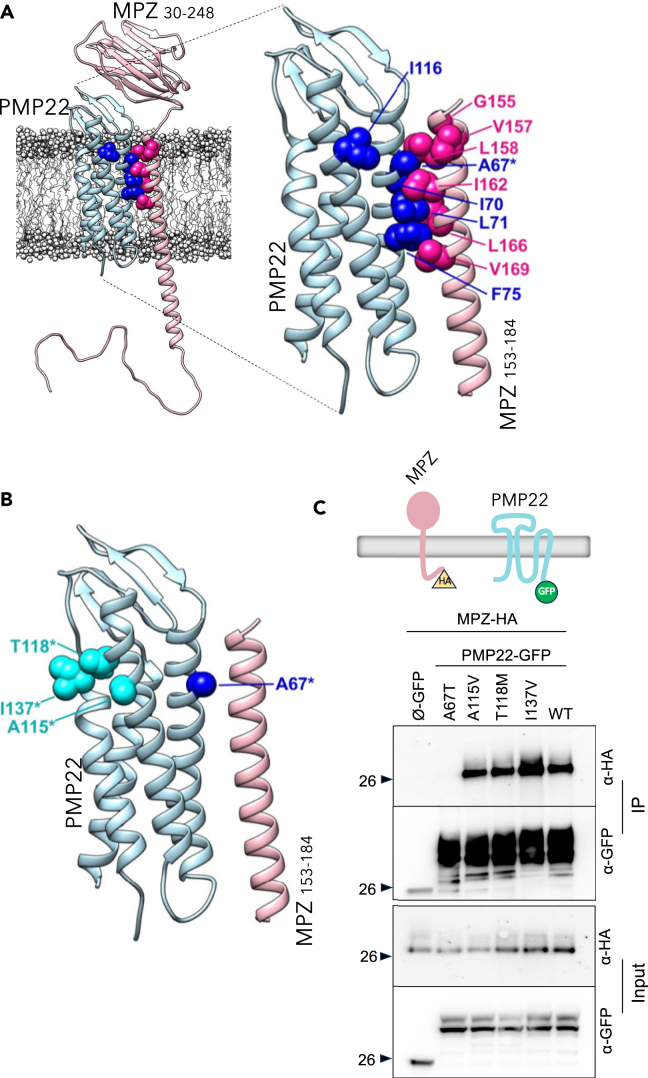
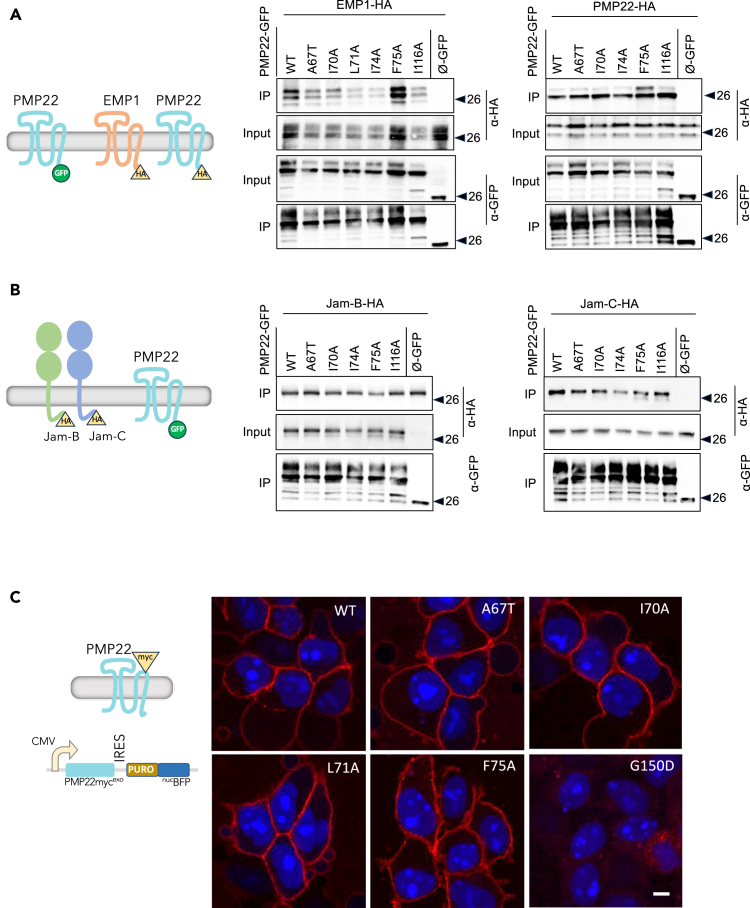
Peripheral Myelin Protein 22 (PMP22) and MPZ are abundant myelin membrane proteins in Schwann cells. The MPZ adhesion protein holds myelin wraps together across the intraperiod line. PMP22 is a tetraspan protein belonging to the Claudin superfamily. Loss of either MPZ or PMP22 causes severe demyelinating Charcot-Marie-Tooth (CMT) peripheral neuropathy, and duplication of PMP22 causes the most common form of CMT, CMT1A. Yet, the molecular functions provided by PMP22 and how its alteration causes CMT are unknown. Here, we find MPZ and PMP22 form a specific complex through interfaces within their transmembrane domains. We also find that the PMP22 A67T patient variant that causes a loss-of-function (hereditary neuropathy with pressure palsies) phenotype maps to this interface, and blocks MPZ association without affecting localization to the plasma membrane or interactions with other proteins. These data define the molecular basis for the MPZ ∼ PMP22 interaction and indicate this complex fulfills an important function in myelinating cells.
Keywords: Biochemistry; Cell biology; Molecular biology; Neuroscience.
© 2024 The Author(s).
Conflict of interest statement
The authors declare there are no competing interests.
Figures



Update of
-
Disrupting the transmembrane domain interface between PMP22 and MPZ causes peripheral neuropathy.bioRxiv [Preprint]. 2024 Sep 11:2023.12.24.573255. doi: 10.1101/2023.12.24.573255. bioRxiv. 2024. Update in: iScience. 2024 Sep 19;27(11):110989. doi: 10.1016/j.isci.2024.110989. PMID: 38187781 Free PMC article. Updated. Preprint.
References
-
- Skre H. Genetic and clinical aspects of Charcot-Marie-Tooth's disease. Clin. Genet. 1974;6:98–118. - PubMed
-
- Fridman V., Bundy B., Reilly M.M., Pareyson D., Bacon C., Burns J., Day J., Feely S., Finkel R.S., Grider T., et al. CMT subtypes and disease burden in patients enrolled in the Inherited Neuropathies Consortium natural history study: a cross-sectional analysis. J. Neurol. Neurosurg. Psychiatry. 2015;86:873–878. doi: 10.1136/jnnp-2014-308826. - DOI - PMC - PubMed
-
- Murphy S.M., Laura M., Fawcett K., Pandraud A., Liu Y.T., Davidson G.L., Rossor A.M., Polke J.M., Castleman V., Manji H., et al. Charcot-Marie-Tooth disease: frequency of genetic subtypes and guidelines for genetic testing. J. Neurol. Neurosurg. Psychiatry. 2012;83:706–710. doi: 10.1136/jnnp-2012-302451. - DOI - PMC - PubMed
-
- Lemke G., Axel R. Isolation and sequence of a cDNA encoding the major structural protein of peripheral myelin. Cell. 1985;40:501–508. - PubMed
-
- Uyemura K., Asou H., Takeda Y. Structure and function of peripheral nerve myelin proteins. Prog. Brain Res. 1995;105:311–318. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
