

Aminoglycoside tolerance in Vibrio cholerae engages translational reprogramming associated with queuosine tRNA modification
- PMID: 39761105
- PMCID: PMC11703503
- DOI: 10.7554/eLife.96317
Aminoglycoside tolerance in Vibrio cholerae engages translational reprogramming associated with queuosine tRNA modification
Abstract
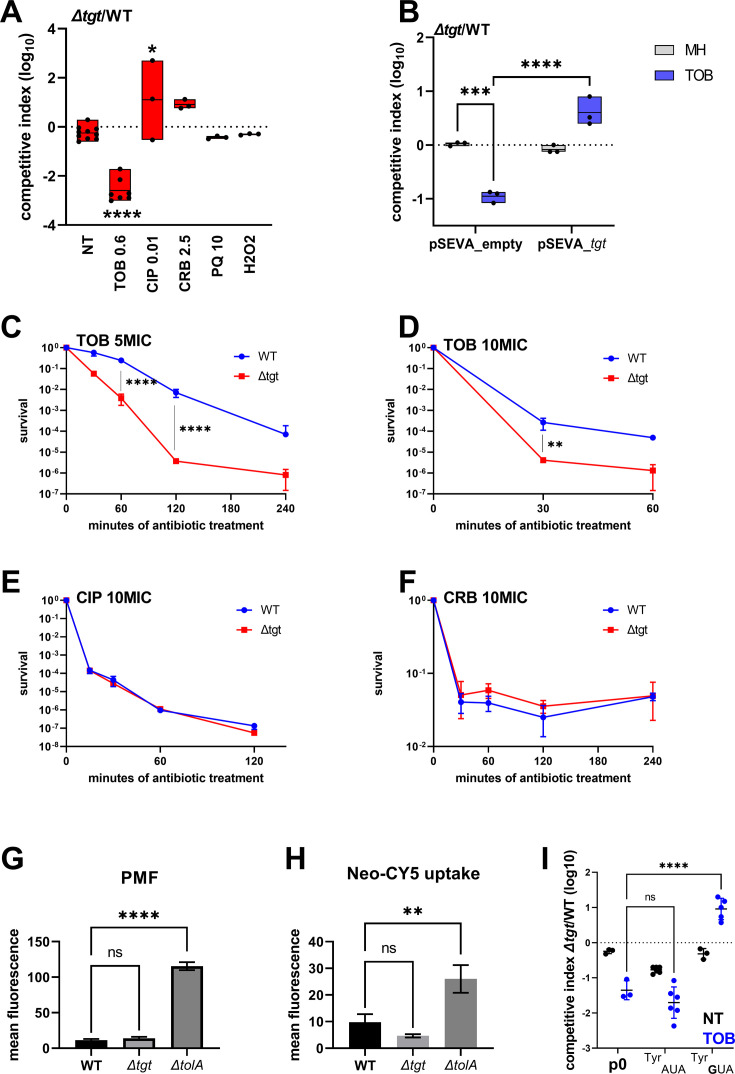
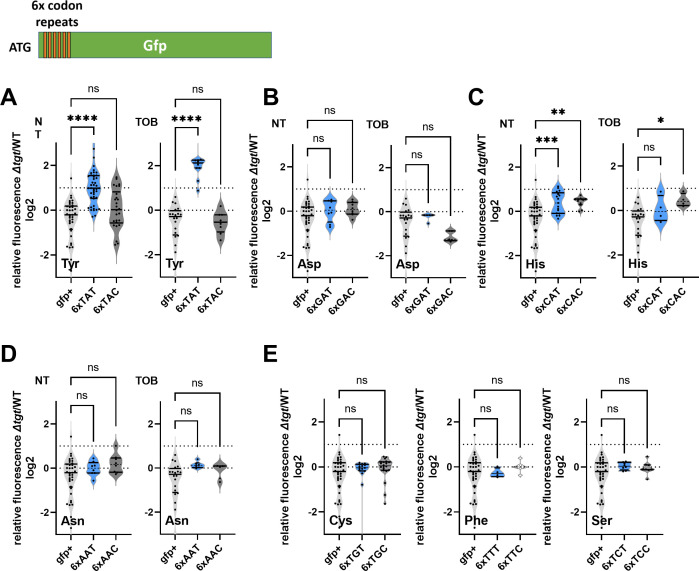
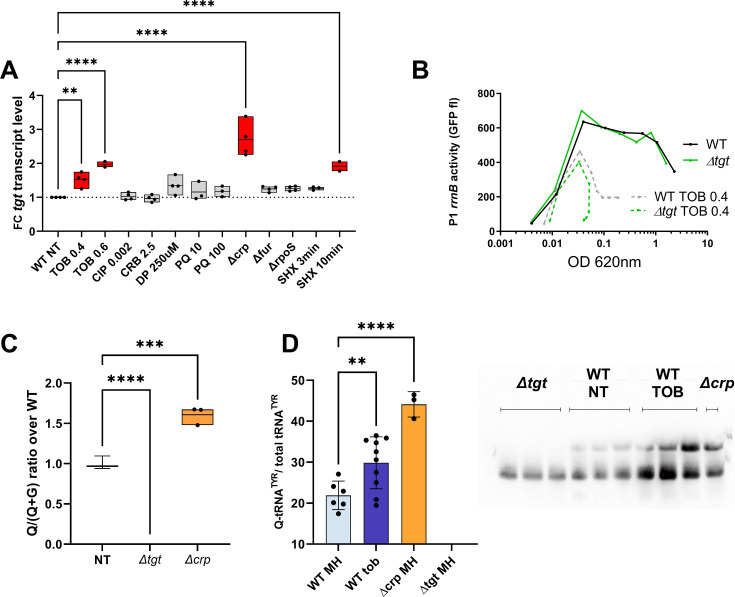
Tgt is the enzyme modifying the guanine (G) in tRNAs with GUN anticodon to queuosine (Q). tgt is required for optimal growth of Vibrio cholerae in the presence of sub-lethal aminoglycoside concentrations. We further explored here the role of the Q34 in the efficiency of codon decoding upon tobramycin exposure. We characterized its impact on the overall bacterial proteome, and elucidated the molecular mechanisms underlying the effects of Q34 modification in antibiotic translational stress response. Using molecular reporters, we showed that Q34 impacts the efficiency of decoding at tyrosine TAT and TAC codons. Proteomics analyses revealed that the anti-SoxR factor RsxA is better translated in the absence of tgt. RsxA displays a codon bias toward tyrosine TAT and overabundance of RsxA leads to decreased expression of genes belonging to SoxR oxidative stress regulon. We also identified conditions that regulate tgt expression. We propose that regulation of Q34 modification in response to environmental cues leads to translational reprogramming of transcripts bearing a biased tyrosine codon usage. In silico analysis further identified candidate genes which could be subject to such translational regulation, among which DNA repair factors. Such transcripts, fitting the definition of modification tunable transcripts, are central in the bacterial response to antibiotics.
Keywords: Vibrio cholerae; antibiotic stress; epitranscriptome; genetics; genomics; infectious disease; microbiology; post-transcriptional regulation; queuosine; tRNA modification.
© 2024, Fruchard, Babosan et al.
Conflict of interest statement
LF, AB, AC, ML, BL, MD, QG, MM, FB, IH, HA, CF, EC, GS, VM, YM, ON, Vd, DM, ZB No competing interests declared
Figures















Update of
- doi: 10.1101/2022.09.26.509455
- doi: 10.7554/eLife.96317.1
- doi: 10.7554/eLife.96317.2
References
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
