

Trichostatin A promotes de novo shoot regeneration from Arabidopsis root explants via a cytokinin related pathway
- PMID: 39762325
- PMCID: PMC11704266
- DOI: 10.1038/s41598-024-84860-9
Trichostatin A promotes de novo shoot regeneration from Arabidopsis root explants via a cytokinin related pathway
Abstract
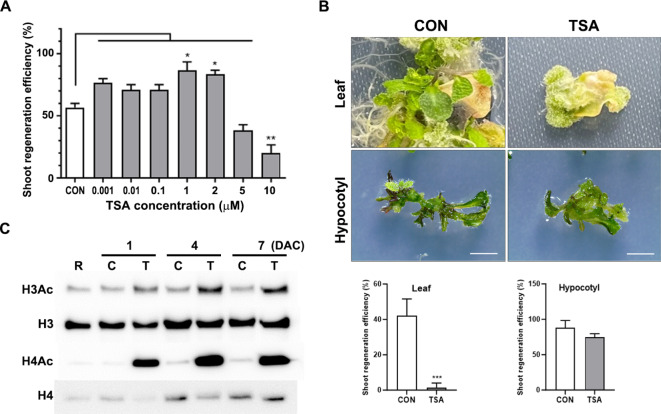
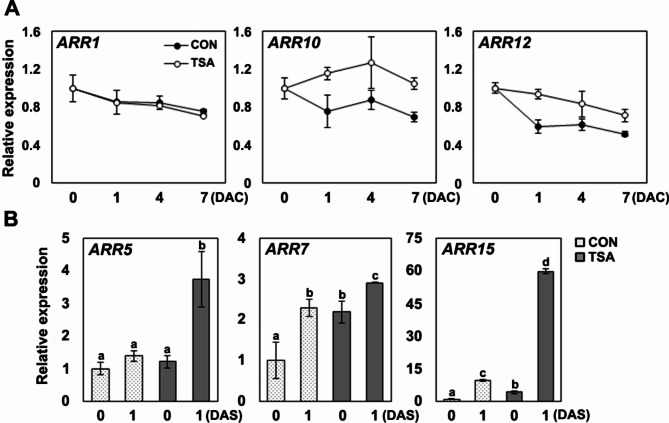
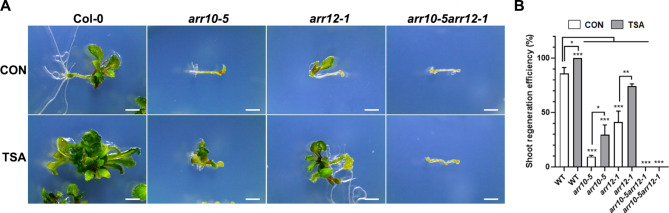
De novo shoot regeneration, characterized by the emergence of adventitious shoots from excised or damaged tissues or organs in vitro, is regulated by the complex interplay between genetic and epigenetic regulatory mechanisms. However, the specific effect of histone deacetylation on shoot regeneration remains poorly understood. This study investigated the effects of trichostatin A (TSA), a histone deacetylase inhibitor, on shoot regeneration in callus derived from root explants. TSA-treated root explants exhibited pronounced callus greening and substantially increasing in multiple shoot formations per callus compared with the control group. Additionally, TSA treatment upregulated shoot apical meristem-specific genes, including WUSCHELL (WUS), RELATED TO AP2.6 L (Rap2.6 L), SHOOT MERISTEMLESS (STM), CUP SHAPED COTYLEDON 2 (CUC2). Notably, TSA treatment enhanced the sensitivity to cytokinins, leading to increase expression of the cytokinin signaling reporter TCS::GFP in the callus. Concomitantly, type-B ARABIDOPSIS RESPONSE REGULATOR (ARR) 10 and 12, which are key regulators of cytokinin signaling, were upregulated in TSA-treated callus, whereas the downstream targets of type-B ARRs, such as ARR5, ARR7, and ARR15, were significantly upregulated during shoot regeneration. Furthermore, mutants deficient in ARR10 and ARR12 showed diminished responsiveness to shoot regenerative capacity, a phenotype that was enhanced by TSA treatment. Our findings underscore the crucial role of histone deacetylation in mediating cytokinin responses and controlling de novo shoot regeneration in plants.
Keywords: Cytokinin response; Histone deacetylation; Shoot regeneration; Trichostatin A.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests. Ethics declarations: Experimental research and field studies on plants, including the collection of plant material, complying with relevant institutional, national, and international guidelines and legislation.
Figures




References
-
- Barton, M. & Poethig, R. S. Formation of the shoot apical meristem in Arabidopsis thaliana—An analysis of development in wild type and in the SHOOT MERISTEMLESS mutant. Development119 (1993).
Publication types
MeSH terms
Substances
Grants and funding
- KGM5382414/KRIBB Initiative Program
- KGM5382414/KRIBB Initiative Program
- KGM5382414/KRIBB Initiative Program
- KGM5282432/KRIBB Initiative Program
- KGM5282432/KRIBB Initiative Program
- KGM5382414/KRIBB Initiative Program
- CAP23051-200/National Research Council of Science & Technology (NST) grant from the Korean government (MSIT)
- CAP23051-200/National Research Council of Science & Technology (NST) grant from the Korean government (MSIT)
- CAP23051-200/National Research Council of Science & Technology (NST) grant from the Korean government (MSIT)
- CAP23051-200/National Research Council of Science & Technology (NST) grant from the Korean government (MSIT)
- CAP23051-200/National Research Council of Science & Technology (NST) grant from the Korean government (MSIT)
- NRF-2020M3A9I4038354/Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government (MSIT)
- NRF-2020M3A9I4038354/Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government (MSIT)
- NRF-2020M3A9I4038354/Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government (MSIT)
- NRF-2020M3A9I4038354/Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government (MSIT)
- NRF-2020M3A9I4038354/Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government (MSIT)
LinkOut - more resources
Full Text Sources
Research Materials
