

This is a preprint.
FAF2 is a bifunctional regulator of peroxisomal homeostasis and saturated lipid responses
- PMID: 39763943
- PMCID: PMC11702540
- DOI: 10.1101/2024.12.12.628015
FAF2 is a bifunctional regulator of peroxisomal homeostasis and saturated lipid responses
Update in
-
FAF2 is a bifunctional regulator of peroxisomal homeostasis and saturated lipid responses.Sci Adv. 2025 Jul 4;11(27):eadu9104. doi: 10.1126/sciadv.adu9104. Epub 2025 Jul 2. Sci Adv. 2025. PMID: 40601736 Free PMC article.
Abstract
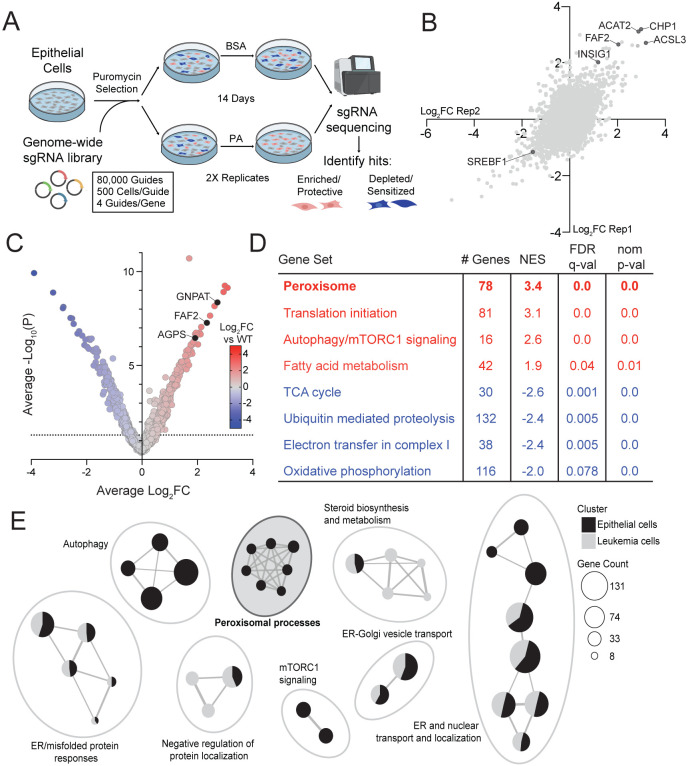
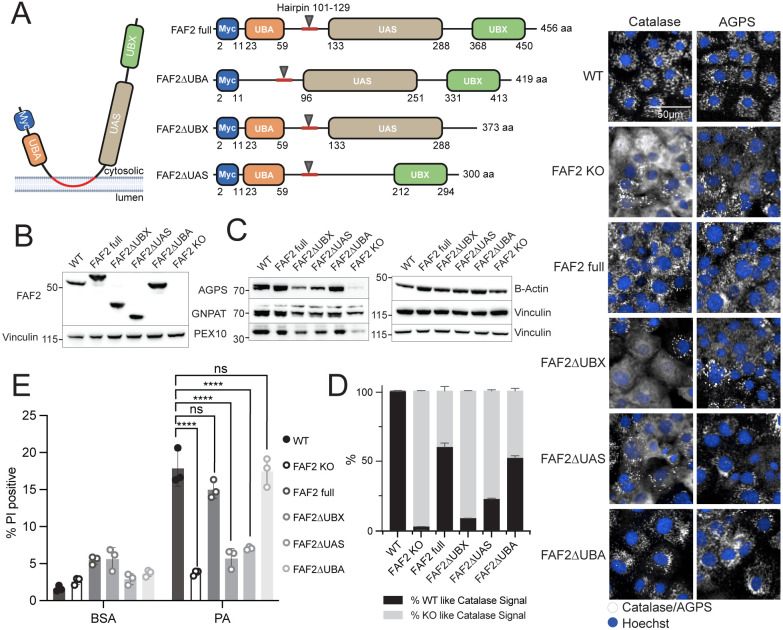
Exposure to saturated fatty acids (SFAs), such as palmitic acid, can lead to cellular metabolic dysfunction known as lipotoxicity. Although canonical adaptive metabolic processes like lipid storage or desaturation are known cellular responses to saturated fat exposure, the link between SFA metabolism and organellar biology remains an area of active inquiry. We performed a genome-wide CRISPR knockout screen in human epithelial cells to identify modulators of SFA toxicity. The screen revealed peroxisomal proteins, especially those that impact ether lipid synthesis, as important regulators of lipotoxicity. We identified Fas-associated factor family member 2 (FAF2) as a critical bifunctional co-regulator of peroxisomal and fatty acid biology. We further uncovered a new biological function for the ubiquitin-regulatory X (UBX) and UAS thioredoxin-like domains of FAF2, demonstrating their requirement for peroxisomal protein abundance and SFA-induced cellular stress. Our work highlights the role of FAF2 in regulating peroxisomal abundance and function, and the peroxisome as a key organelle in the cellular response to SFAs.
Figures



References
-
- Weinberg J. M., Lipotoxicity. Kidney Int. 70, 1560–1566 (2006). - PubMed
-
- Piccolis M., Bond L. M., Kampmann M., Pulimeno P., Chitraju C., Jayson C. B. K., Vaites L. P., Boland S., Lai Z. W., Gabriel K. R., Elliott S. D., Paulo J. A., Harper J. W., Weissman J. S., Walther T. C., Farese R. V. Jr, Probing the global cellular responses to lipotoxicity caused by saturated fatty acids. Mol. Cell 74, 32–44.e8 (2019). - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous