

This is a preprint.
Tax1bp1 enhances bacterial virulence and promotes inflammatory responses during Mycobacterium tuberculosis infection of alveolar macrophages
- PMID: 39763950
- PMCID: PMC11702572
- DOI: 10.1101/2024.12.16.628616
Tax1bp1 enhances bacterial virulence and promotes inflammatory responses during Mycobacterium tuberculosis infection of alveolar macrophages
Abstract
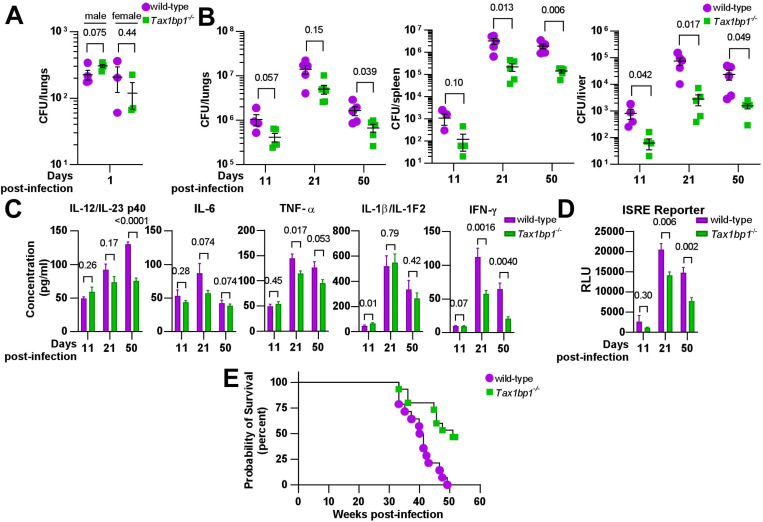
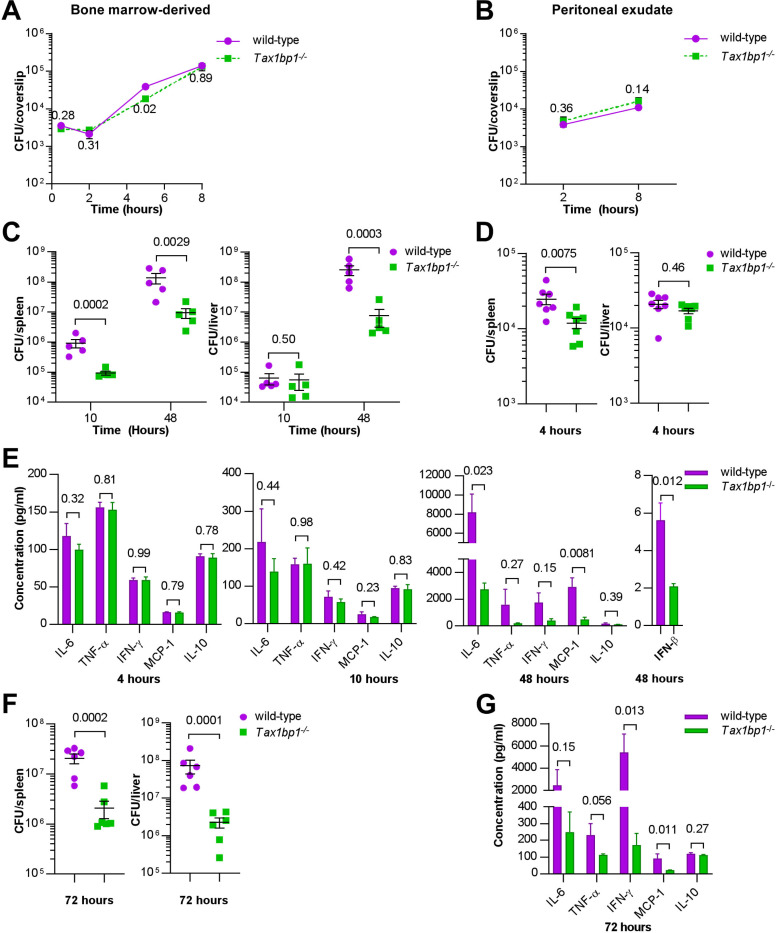
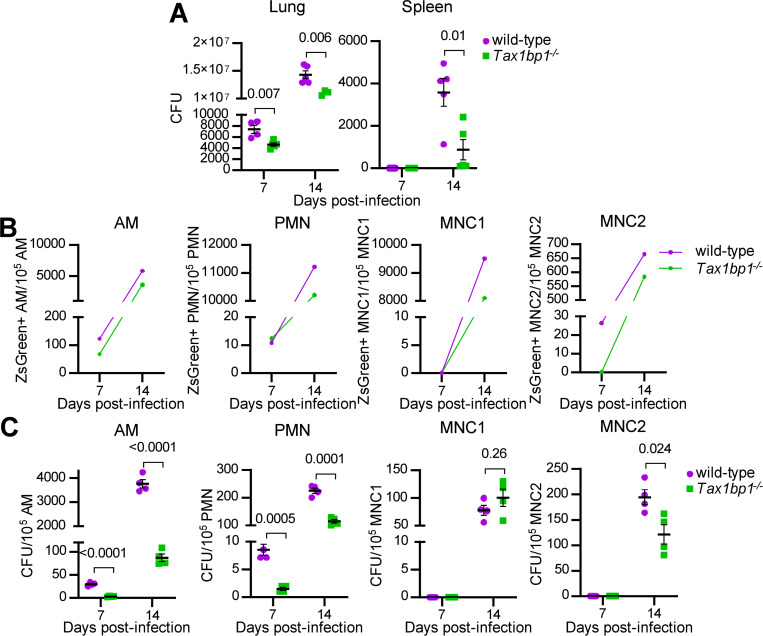
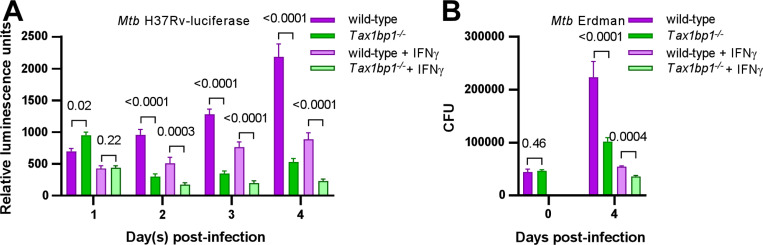
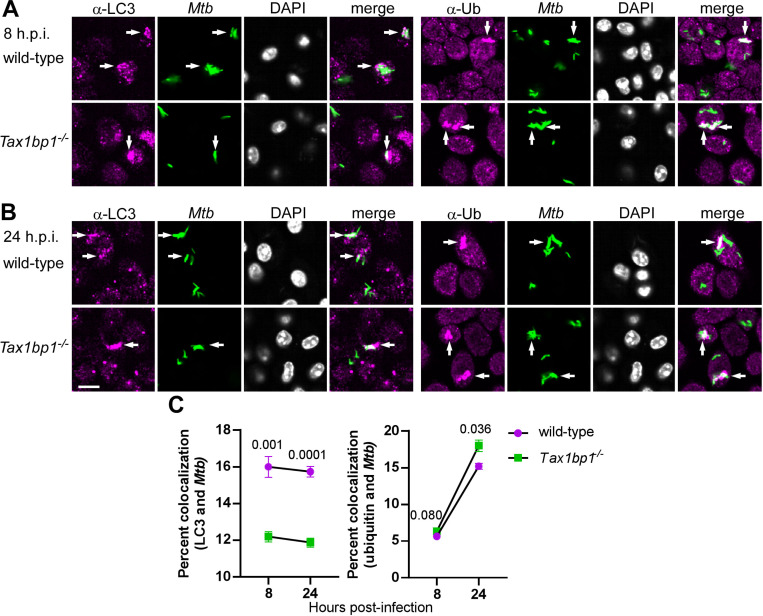
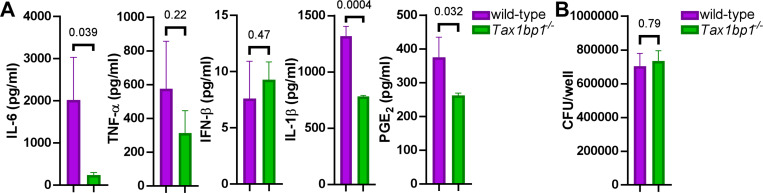
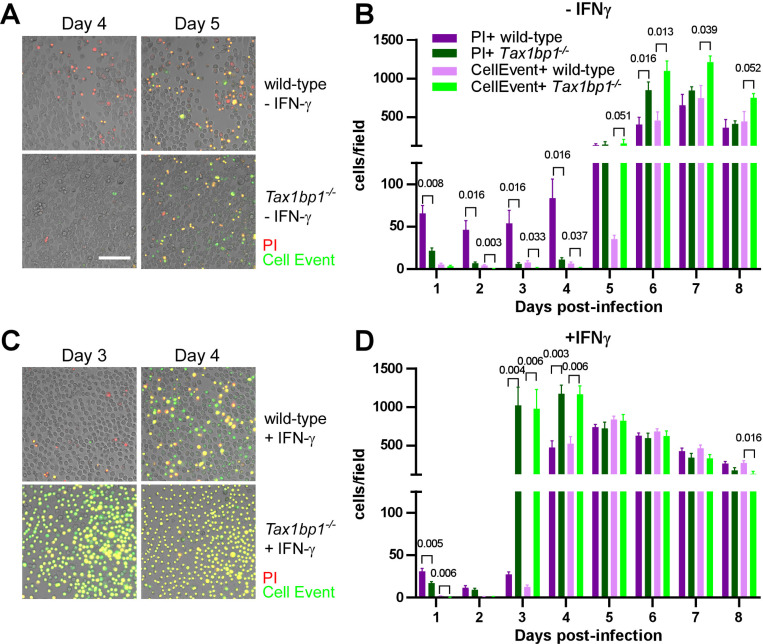
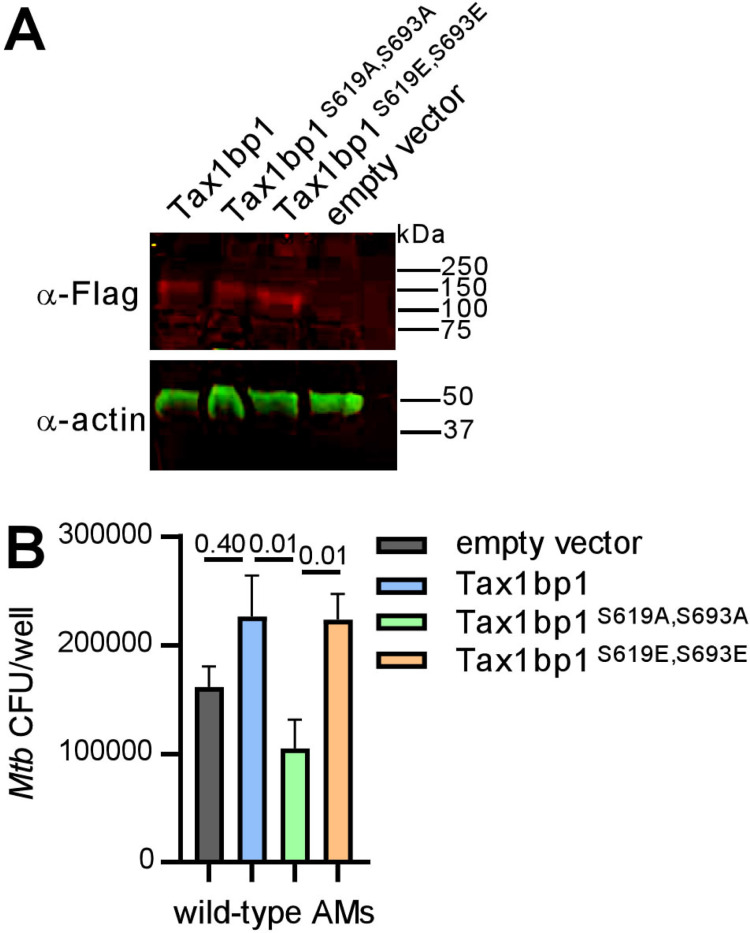
Crosstalk between autophagy, host cell death, and inflammatory host responses to bacterial pathogens enables effective innate immune responses that limit bacterial growth while minimizing coincidental host damage. Mycobacterium tuberculosis (Mtb) thwarts innate immune defense mechanisms in alveolar macrophages (AMs) during the initial stages of infection and in recruited bone marrow-derived cells during later stages of infection. However, how protective inflammatory responses are achieved during Mtb infection and the variation of the response in different macrophage subtypes remain obscure. Here, we show that the autophagy receptor Tax1bp1 plays a critical role in enhancing inflammatory cytokine production and increasing the susceptibility of mice to Mtb infection. Surprisingly, although Tax1bp1 restricts Mtb growth during infection of bone marrow-derived macrophages (BMDMs) (Budzik et al. 2020) and terminates cytokine production in response to cytokine stimulation or viral infection, Tax1bp1 instead promotes Mtb growth in AMs, neutrophils, and a subset of recruited monocyte-derived cells from the bone marrow. Tax1bp1 also leads to increases in bacterial growth and inflammatory responses during infection of mice with Listeria monocytogenes, an intracellular pathogen that is not effectively targeted to canonical autophagy. In Mtb-infected AMs but not BMDMs, Tax1bp1 enhances necrotic-like cell death early after infection, reprogramming the mode of host cell death to favor Mtb replication in AMs. Tax1bp1's impact on host cell death is a mechanism that explains Tax1bp1's cell type-specific role in the control of Mtb growth. Similar to Tax1bp1-deficiency in AMs, the expression of phosphosite-deficient Tax1bp1 restricts Mtb growth. Together, these results show that Tax1bp1 plays a crucial role in linking the regulation of autophagy, cell death, and pro-inflammatory host responses and enhancing susceptibility to bacterial infection.
Figures

References
-
- World Health Organization. Global Tuberculosis Report 2023. Geneva: World Health Organization; 2023. Nov.
Publication types
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources