

Metabolic reprogramming of macrophages by PKM2 promotes IL-10 production via adenosine
- PMID: 39772395
- PMCID: PMC11781862
- DOI: 10.1016/j.celrep.2024.115172
Metabolic reprogramming of macrophages by PKM2 promotes IL-10 production via adenosine
Abstract
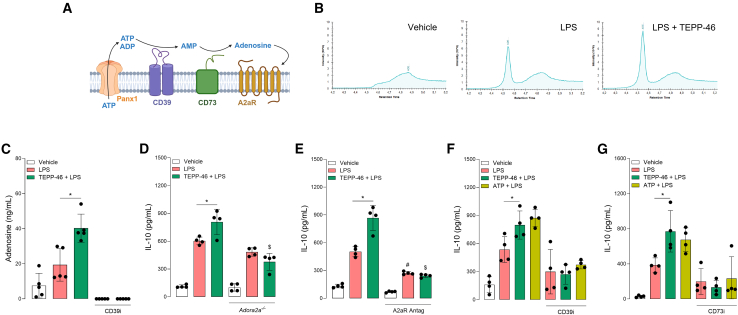
Macrophages play a crucial role in immune responses and undergo metabolic reprogramming to fulfill their functions. The tetramerization of the glycolytic enzyme pyruvate kinase M2 (PKM2) induces the production of the anti-inflammatory cytokine interleukin (IL)-10 in vivo, but the underlying mechanism remains elusive. Here, we report that PKM2 activation with the pharmacological agent TEPP-46 increases IL-10 production in LPS-activated macrophages by metabolic reprogramming, leading to the production and release of ATP from glycolysis. The effect of TEPP-46 is abolished in PKM2-deficient macrophages. Extracellular ATP is converted into adenosine by ectonucleotidases that activate adenosine receptor A2a (A2aR) to enhance IL-10 production. Interestingly, IL-10 production induced by PKM2 activation is associated with improved mitochondrial health. Our results identify adenosine derived from glycolytic ATP as a driver of IL-10 production, highlighting the role of tetrameric PKM2 in regulating glycolysis to promote IL-10 production.
Keywords: CP: Immunology; CP: Metabolism; IL-10; PKM2; adenosine; macrophage; mitochondria dynamics.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




References
-
- Freemerman A.J., Johnson A.R., Sacks G.N., Milner J.J., Kirk E.L., Troester M.A., Macintyre A.N., Goraksha-Hicks P., Rathmell J.C., Makowski L. Metabolic reprogramming of macrophages: glucose transporter 1 (GLUT1)-mediated glucose metabolism drives a proinflammatory phenotype. J. Biol. Chem. 2014;289:7884–7896. doi: 10.1074/jbc.M113.522037. - DOI - PMC - PubMed
-
- Jha A.K., Huang S.C.C., Sergushichev A., Lampropoulou V., Ivanova Y., Loginicheva E., Chmielewski K., Stewart K.M., Ashall J., Everts B., et al. Network integration of parallel metabolic and transcriptional data reveals metabolic modules that regulate macrophage polarization. Immunity. 2015;42:419–430. doi: 10.1016/j.immuni.2015.02.005. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
