

BK channels mediate a presynaptic form of mGluR-LTD in the neonatal hippocampus
- PMID: 39773031
- PMCID: PMC11745352
- DOI: 10.1073/pnas.2411506122
BK channels mediate a presynaptic form of mGluR-LTD in the neonatal hippocampus
Abstract
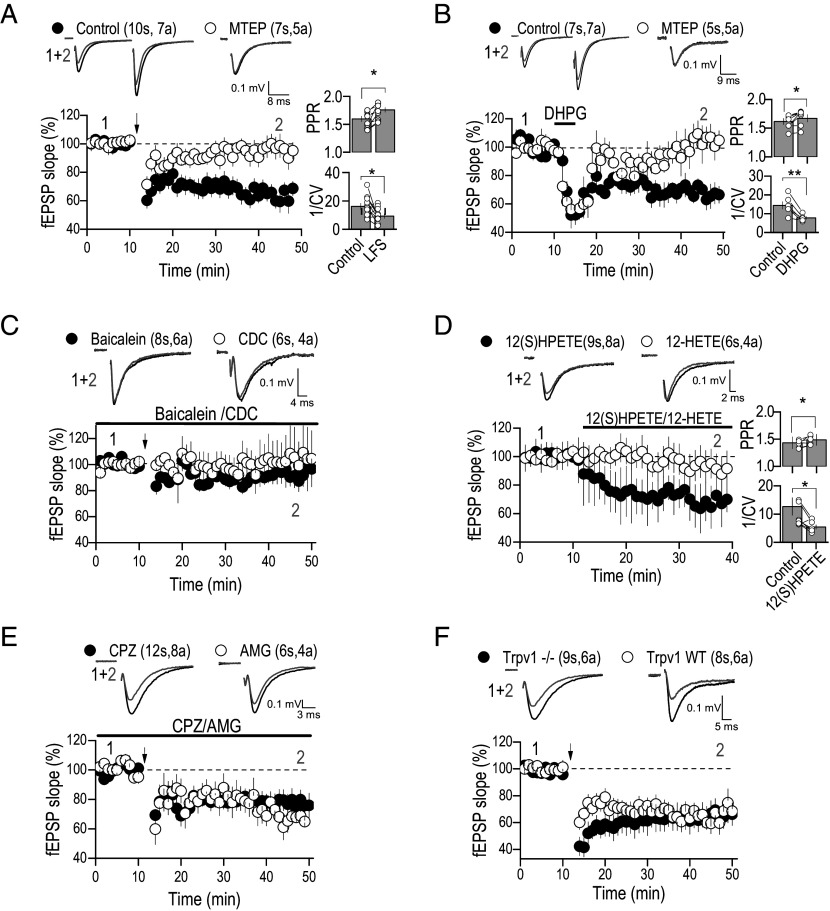
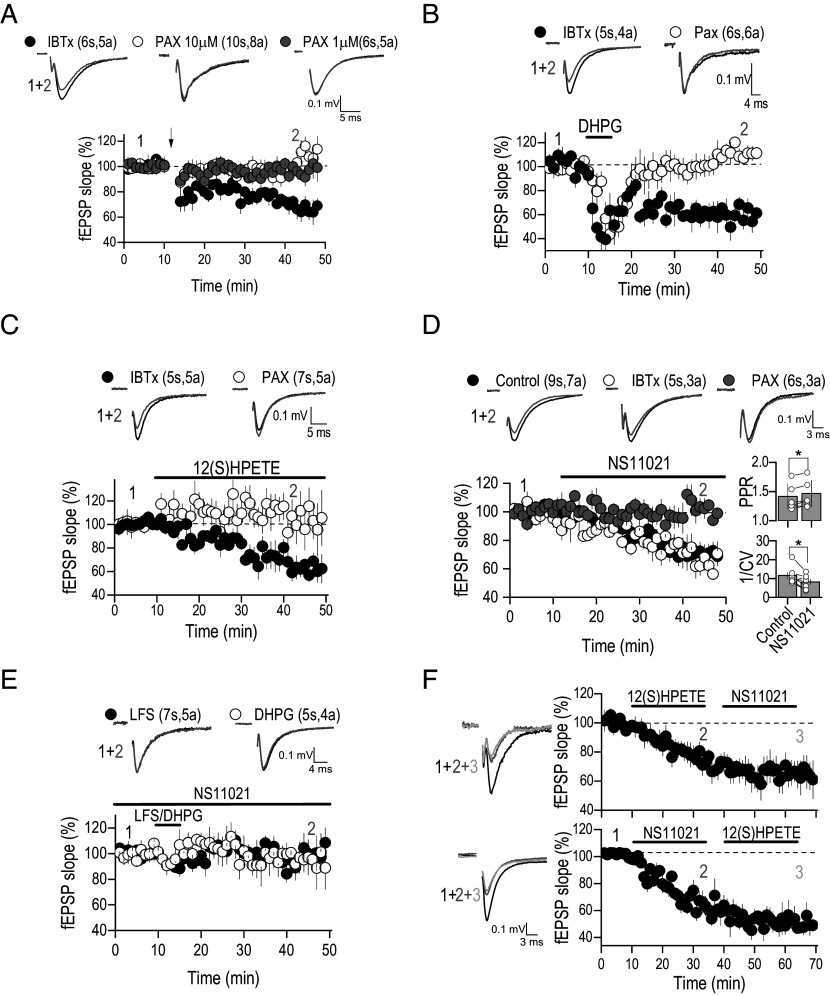
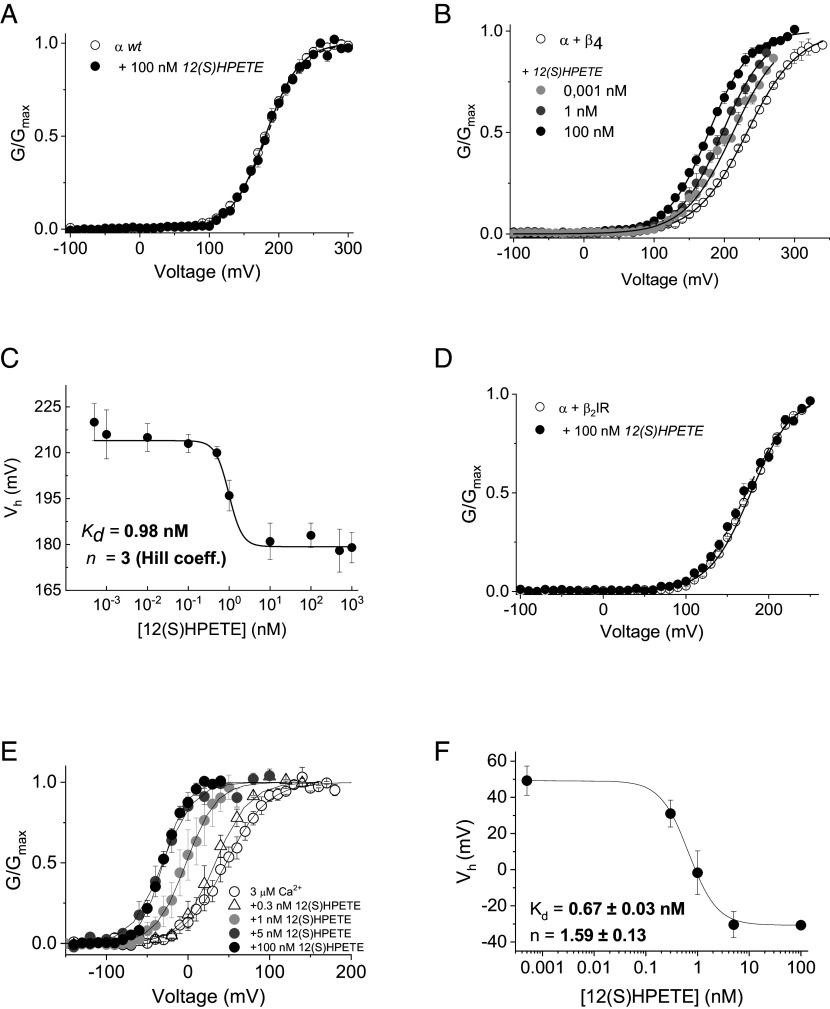
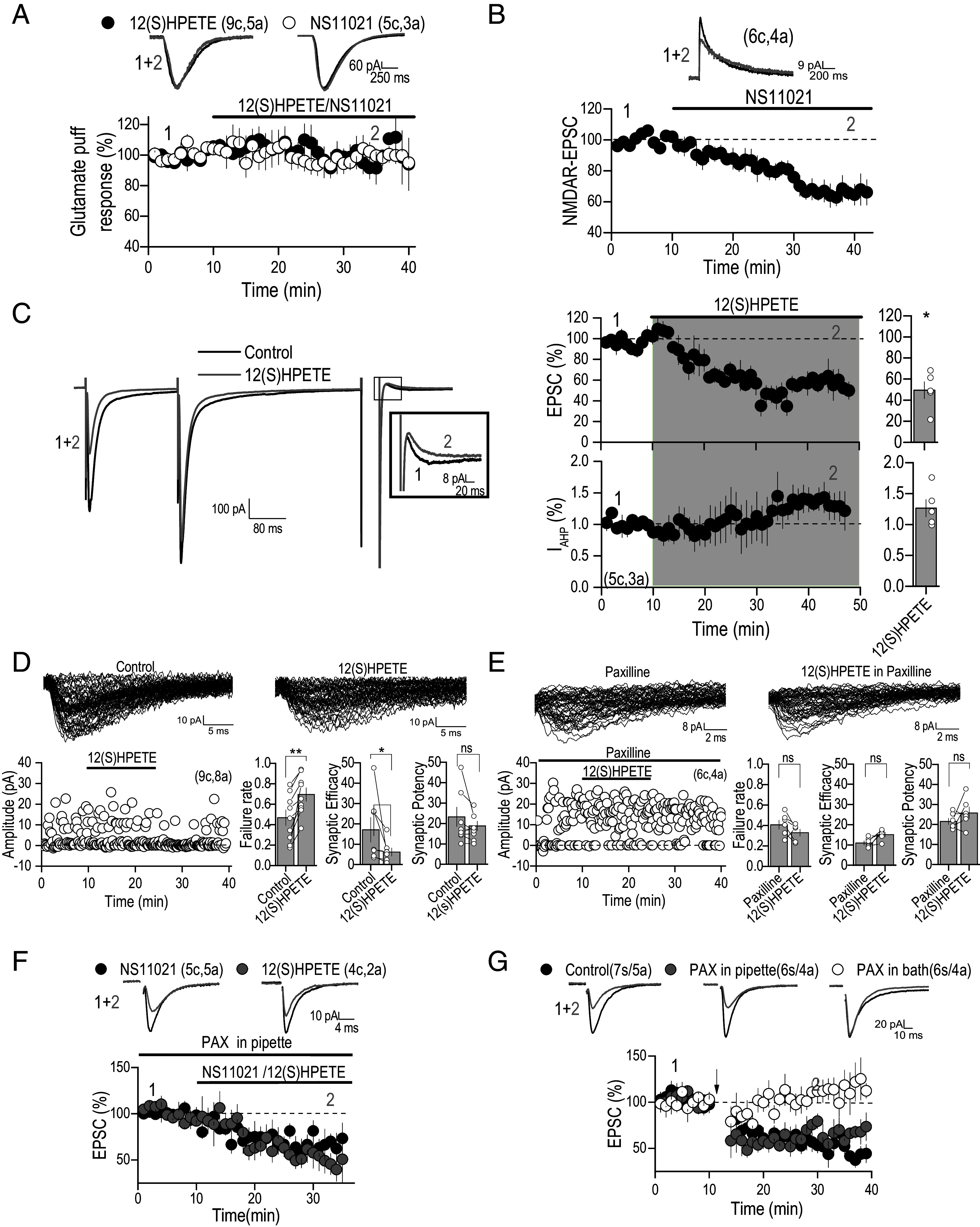
BK channels can control neuronal function, but their functional relevance in activity-dependent changes of synaptic function remains elusive. Here, we report that repetitive low-frequency stimulation activates BK channels through 12(S)HPETE, an arachidonic acid metabolite, produced downstream of postsynaptic metabotropic glutamate receptors (mGluRs) to trigger long-term depression (LTD) at CA3-CA1 synapses in hippocampal slices from P7-P10 mice. Activation of BK channels is subunit specific, as paxilline but not iberiotoxin blocked mGluR-LTD. Also, 12(S)HPETE does not change the electrophysiological properties of the BK channel when the BKα subunit is expressed alone but increases the channel open probability when the BKα is coexpressed with the β4-subunit. Our findings reveal an interaction between 12(S)HPETE and BK channels to regulate synaptic strength at central synapses and increase our understanding of the mechanisms underlying mGluR-LTD in the neonatal hippocampus that likely contribute to circuit maturation necessary for learning.
Keywords: BK channels; hippocampus; retrograde signaling; synaptic plasticity.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
Similar articles
-
12-lipoxygenase metabolites of arachidonic acid mediate metabotropic glutamate receptor-dependent long-term depression at hippocampal CA3-CA1 synapses.J Neurosci. 2003 Dec 10;23(36):11427-35. doi: 10.1523/JNEUROSCI.23-36-11427.2003. J Neurosci. 2003. PMID: 14673007 Free PMC article.
-
Endocannabinoid-mediated long-term depression of afferent excitatory synapses in hippocampal pyramidal cells and GABAergic interneurons.J Neurosci. 2012 Oct 10;32(41):14448-63. doi: 10.1523/JNEUROSCI.1676-12.2012. J Neurosci. 2012. PMID: 23055515 Free PMC article.
-
Loss of anoctamin 1 reveals a subtle role for BK channels in lymphatic muscle action potentials.J Physiol. 2024 Jul;602(14):3351-3373. doi: 10.1113/JP285459. Epub 2024 May 5. J Physiol. 2024. PMID: 38704841 Free PMC article.
-
Short term plasticity at hippocampal mossy fiber synapses.Neuroscience. 2025 Jul 10;578:105-117. doi: 10.1016/j.neuroscience.2024.09.044. Epub 2024 Sep 26. Neuroscience. 2025. PMID: 39332701 Review.
-
Maternal and neonatal outcomes of elective induction of labor.Evid Rep Technol Assess (Full Rep). 2009 Mar;(176):1-257. Evid Rep Technol Assess (Full Rep). 2009. PMID: 19408970 Free PMC article.
References
-
- Bolshakov V. Y., Siegelbaum S. A., Postsynaptic induction and presynaptic expression of hippocampal long-term depression. Science 264, 1148–1152 (1994). - PubMed
-
- Bolshakov V. Y., Siegelbaum S. A., Regulation of hippocampal transmitter release during development and long-term potentiation. Science 269, 1730–1734 (1995). - PubMed
MeSH terms
Substances
Grants and funding
- 1201848/ANID | Fondo Nacional de Desarrollo Científico y Tecnológico (FONDECYT)
- P09-022F/Agencia Nacional de Investigación y Desarrollo (ANID)
- 1230265/ANID | Fondo Nacional de Desarrollo Científico y Tecnológico (FONDECYT)
- R01 GM030376/GM/NIGMS NIH HHS/United States
- 21202136/Agencia Nacional de Investigación y Desarrollo (ANID)
LinkOut - more resources
Full Text Sources
Miscellaneous
