

VCP regulates early tau seed amplification via specific cofactors
- PMID: 39773263
- PMCID: PMC11707990
- DOI: 10.1186/s13024-024-00783-z
VCP regulates early tau seed amplification via specific cofactors
Abstract
Background: Neurodegenerative tauopathies may progress based on seeding by pathological tau assemblies, whereby an aggregate is released from one cell, gains entry to an adjacent or connected cell, and serves as a specific template for its own replication in the cytoplasm. Seeding into the complex cytoplasmic milieu happens within hours, implying the existence of unknown factors that regulate this process.
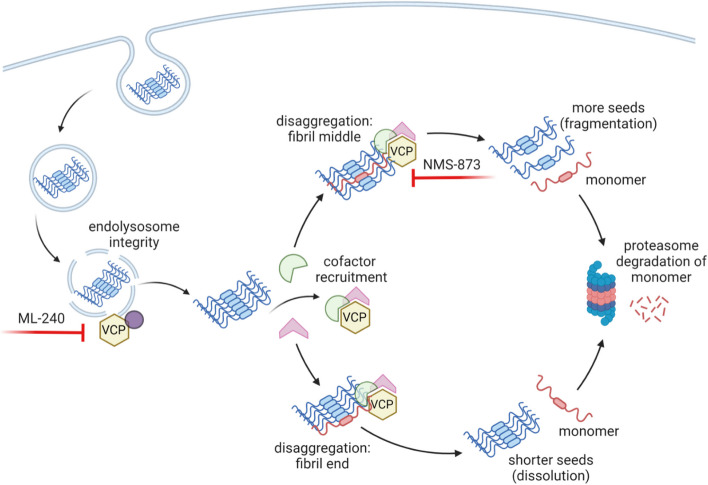
Methods: We used proximity labeling to identify proteins that control seed amplification within 5 h of seed exposure. We fused split-APEX2 to the C-terminus of tau repeat domain (RD) to reconstitute peroxidase activity 5 h after seeded intracellular tau aggregation. Valosin containing protein (VCP/p97) was the top hit. VCP harbors dominant mutations that underlie two neurodegenerative diseases, multisystem proteinopathy and vacuolar tauopathy, but its mechanistic role is unclear. We used immortalized cells and human neurons to study the effects of VCP on tau seeding. We exposed cells to fibrils or brain homogenates in cell culture media and measured effects on uptake and induction of intracellular tau aggregation following various genetic and pharmacological manipulations of VCP.
Results: VCP knockdown reduced tau seeding. Chemical inhibitors had opposing effects on seeding in HEK293T tau biosensor cells and human neurons: ML-240 increased seeding efficiency, whereas NMS-873 decreased it. The inhibitors only functioned when administered within 8 h of seed exposure, indicating a role for VCP early in seed processing. We screened 30 VCP co-factors in HEK293T biosensor cells by genetic knockout or knockdown. Reduction of ATXN3, NSFL1C, UBE4B, NGLY1, and OTUB1 decreased tau seeding, as did NPLOC4, which also uniquely increased soluble tau levels. By contrast, reduction of FAF2 increased tau seeding.
Conclusions: Divergent effects on tau seeding of chemical inhibitors and cofactor reduction indicate that VCP regulates this process. This is consistent with a cytoplasmic processing complex centered on VCP that directs seeds acutely towards degradation vs. amplification.
Keywords: APEX2; Cofactors; Disaggregase; Seeding; Tau; VCP; p97.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Consent for publication: The authors give consent for publication. Competing interests: The authors declare no competing interests.
Figures








Update of
-
VCP regulates early tau seed amplification via specific cofactors.Res Sq [Preprint]. 2024 May 22:rs.3.rs-4307848. doi: 10.21203/rs.3.rs-4307848/v1. Res Sq. 2024. Update in: Mol Neurodegener. 2025 Jan 7;20(1):2. doi: 10.1186/s13024-024-00783-z. PMID: 38826306 Free PMC article. Updated. Preprint.
References
-
- Lee VM, Goedert M, Trojanowski JQ. Neurodegenerative tauopathies. Annu Rev Neurosci. 2001;24:1121–59. - PubMed
-
- Braak H, Braak E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991;82(4):239–59. - PubMed
-
- Hoenig MC, Bischof GN, Seemiller J, Hammes J, Kukolja J, Onur ÖA, et al. Networks of tau distribution in Alzheimer’s disease. Brain. 2018Feb 1;141(2):568–81. - PubMed
-
- Ramirez DMO, Whitesell JD, Bhagwat N, Thomas TL, Ajay AD, Nawaby A, et al. Endogenous pathology in tauopathy mice progresses via brain networks [Internet]. bioRxiv; 2023 [cited 2023 Jul 19]. p. 2023.05.23.541792. Available from: https://www.biorxiv.org/content/10.1101/2023.05.23.541792v1
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
