

Identification and monitoring of cell heterogeneity from plasmid recombination during limonene production
- PMID: 39775940
- PMCID: PMC11706860
- DOI: 10.1007/s00253-024-13273-5
Identification and monitoring of cell heterogeneity from plasmid recombination during limonene production
Abstract
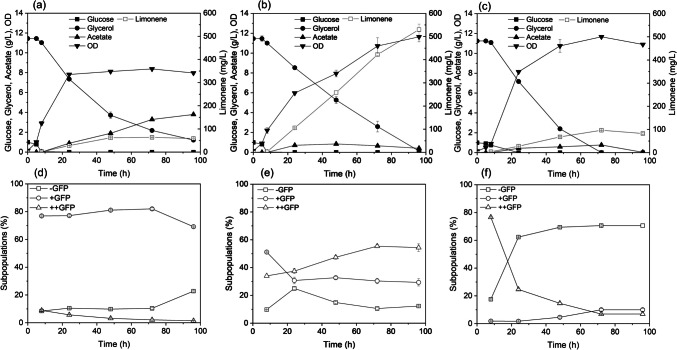
Detecting alterations in plasmid structures is often performed using conventional molecular biology. However, these methods are laborious and time-consuming for studying the conditions inducing these mutations, which prevent real-time access to cell heterogeneity during bioproduction. In this work, we propose combining both flow cytometry and fluorescence-activated cell sorting, integrated with mechanistic modelling to study conditions that lead to plasmid recombination using a limonene-producing microbial system as a case study. A gene encoding GFP was introduced downstream of the key enzymes involved in limonene biosynthesis to enable real-time kinetics monitoring and the identification of cell heterogeneity according to microscopic and flow cytometric analyses. Three different plasmid configurations (one correct and two incorrect) were identified through cell sorting based on subpopulations expressing different levels of GFP at 10 and 50 µM IPTG. Higher limonene production (530 mg/L) and lower subpopulation proportion carrying the incorrect plasmid (12%) were observed for 10 µM IPTG compared to 50 µM IPTG (96 mg/L limonene and more than 70% of cell population carrying the incorrect plasmid, respectively) in 100 mL production culture. We also managed to derive exploratory hypotheses regarding the plasmid recombination region using the model and successfully validated them experimentally. Additionally, the results also showed that limonene production was proportional to GFP fluorescence intensity. This correlation could serve as an alternative to using biosensors for a high-throughput screening process. The developed method enables rapid identification of plasmid recombination at single-cell level and correlates the heterogeneity with bioproduction performance. KEY POINTS: • Strategy to study plasmid recombination during bioproduction. • Different plasmid structures can be identified and monitored by flow cytometry. • Mathematical modelling suggests specific alterations in plasmid structures.
Keywords: Cell heterogeneity; Flow cytometry; Fluorescence-activated cell sorting; Limonene; Mathematical modelling; Plasmid recombination.
© 2025. The Author(s).
Conflict of interest statement
Declarations. This article does not contain any studies with animals performed by any of the authors. Conflict of interest: The authors declare no competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
