

Calcium-sensing receptor- and ADAM10-mediated klotho shedding is regulated by tetraspanin 5
- PMID: 39777735
- PMCID: PMC11931982
- DOI: 10.1002/1873-3468.15078
Calcium-sensing receptor- and ADAM10-mediated klotho shedding is regulated by tetraspanin 5
Abstract
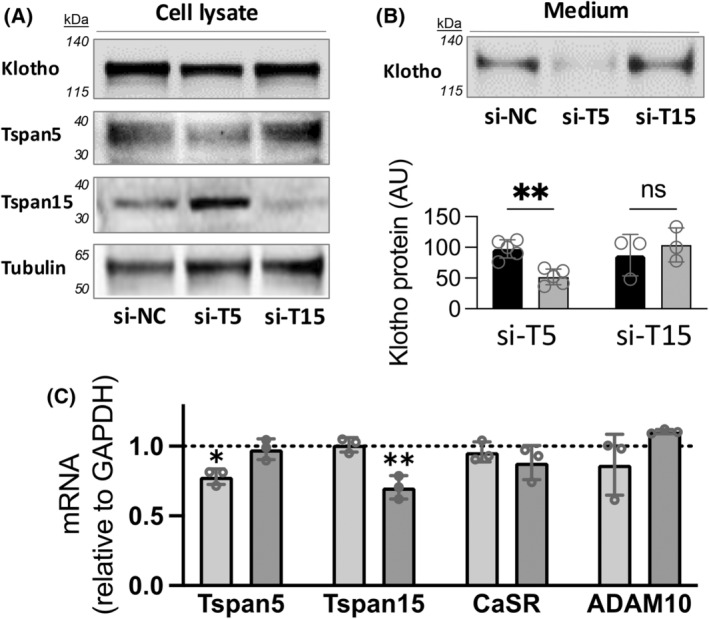
Soluble, circulating Klotho (sKlotho) is essential for normal health and renal function. sKlotho is shed from the renal distal convoluted tubule (DCT), its primary source, via enzymatic cleavage. However, the physiologic mechanisms that control sKlotho production, trafficking, and shedding are not fully defined. We previously found that the G protein-coupled calcium-sensing receptor (CaSR) co-localizes with membrane-bound αKlotho and the disintegrin/metalloprotease ADAM10 in the DCT and controls sKlotho in response to CaSR ligands and pHo by activating ADAM10. Here, we advance understanding of this process by showing that tetraspanin 5 (Tspan5), a scaffolding and chaperone protein, contributes to the cell surface expression and specificity of a protein complex that includes Tspan5, ADAM10, Klotho, and CaSR. These results support a model of multiprotein complexes that confer signaling specificity beyond CaSR on G protein-coupled processes. Impact statement Systemic circulating sKlotho is a determinant for normal physiology. Studies of knockout animals established its role as an anti-aging protein. The regulatory mechanisms for Klotho production and secretion are largely unknown. We report that Tspan 5 contributes to CaSR- and ADAM10-dependent Klotho shedding from the kidney, its primary source.
Keywords: ADAM10; Klotho; Tspan; calcim‐sensing receptor; distal convoluted tubule.
© 2025 The Author(s). FEBS Letters published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Figures



Similar articles
-
Physiologic Regulation of Systemic Klotho Levels by Renal CaSR Signaling in Response to CaSR Ligands and pH o.J Am Soc Nephrol. 2021 Dec 1;32(12):3051-3065. doi: 10.1681/ASN.2021020276. Epub 2021 Dec 1. J Am Soc Nephrol. 2021. PMID: 34551996 Free PMC article.
-
The tetraspanin Tspan15 is an essential subunit of an ADAM10 scissor complex.J Biol Chem. 2020 Sep 4;295(36):12822-12839. doi: 10.1074/jbc.RA120.012601. Epub 2020 Feb 28. J Biol Chem. 2020. PMID: 32111735 Free PMC article.
-
New insights into the tetraspanin Tspan5 using novel monoclonal antibodies.J Biol Chem. 2017 Jun 9;292(23):9551-9566. doi: 10.1074/jbc.M116.765669. Epub 2017 Apr 20. J Biol Chem. 2017. PMID: 28428248 Free PMC article.
-
Regulation of the trafficking and the function of the metalloprotease ADAM10 by tetraspanins.Biochem Soc Trans. 2017 Aug 15;45(4):937-44. doi: 10.1042/BST20160296. Epub 2017 Jul 7. Biochem Soc Trans. 2017. PMID: 28687716 Review.
-
Regulation of ADAM10 by the TspanC8 Family of Tetraspanins and Their Therapeutic Potential.Int J Mol Sci. 2021 Jun 23;22(13):6707. doi: 10.3390/ijms22136707. Int J Mol Sci. 2021. PMID: 34201472 Free PMC article. Review.
References
-
- Sato T, Courbebaisse M, Ide N, Fan Y, Hanai JI, Kaludjerovic J, Densmore MJ, Yuan Q, Toka HR, Pollak MR et al. (2017) Parathyroid hormone controls paracellular Ca(2+) transport in the thick ascending limb by regulating the tight‐junction protein Claudin14. Proc Natl Acad Sci USA 114, E3344–E3353. - PMC - PubMed
-
- Huang C, Sindic A, Hill CE, Hujer KM, Chan KW, Sassen M, Wu Z, Kurachi Y, Nielsen S, Romero MF et al. (2007) Interaction of the Ca‐sensing receptor with the inwardly‐rectifying potassium channels Kir4.1 and Kir4.2 results in inhibition of channel function. Am J Physiol Renal Physiol 292, F1073–F1081. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
