

Auditory evoked-potential abnormalities in a mouse model of 22q11.2 Deletion Syndrome and their interactions with hearing impairment
- PMID: 39779687
- PMCID: PMC11711659
- DOI: 10.1038/s41398-024-03218-x
Auditory evoked-potential abnormalities in a mouse model of 22q11.2 Deletion Syndrome and their interactions with hearing impairment
Abstract
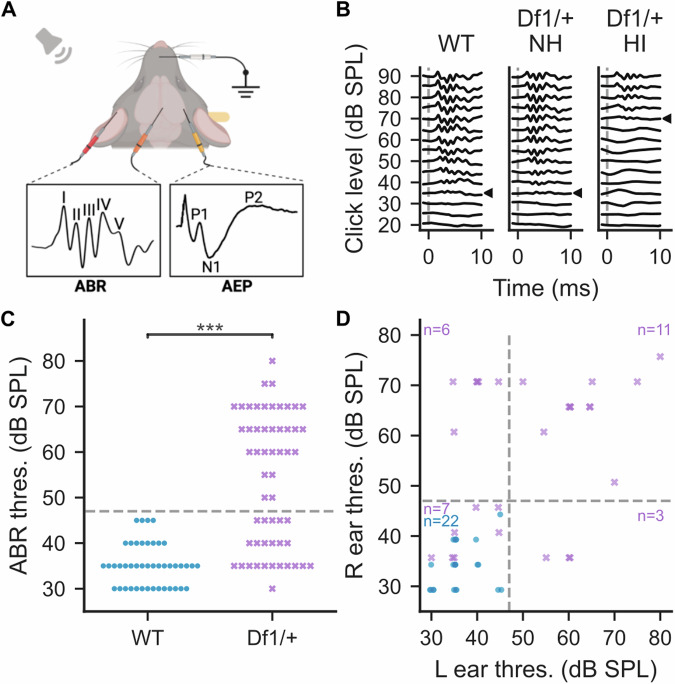
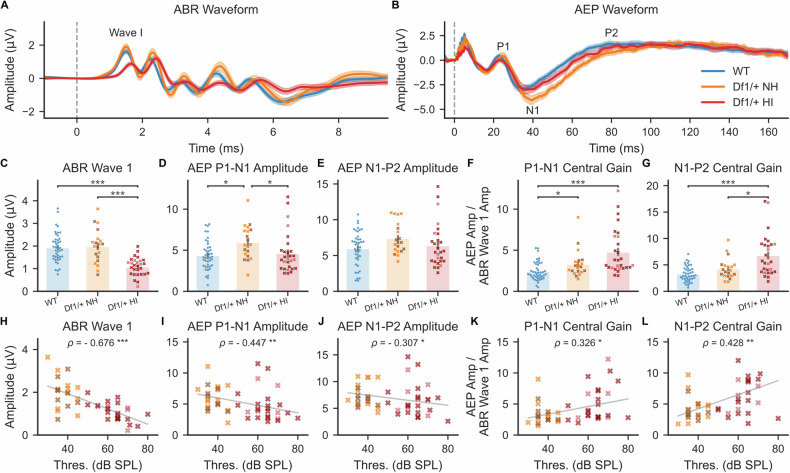
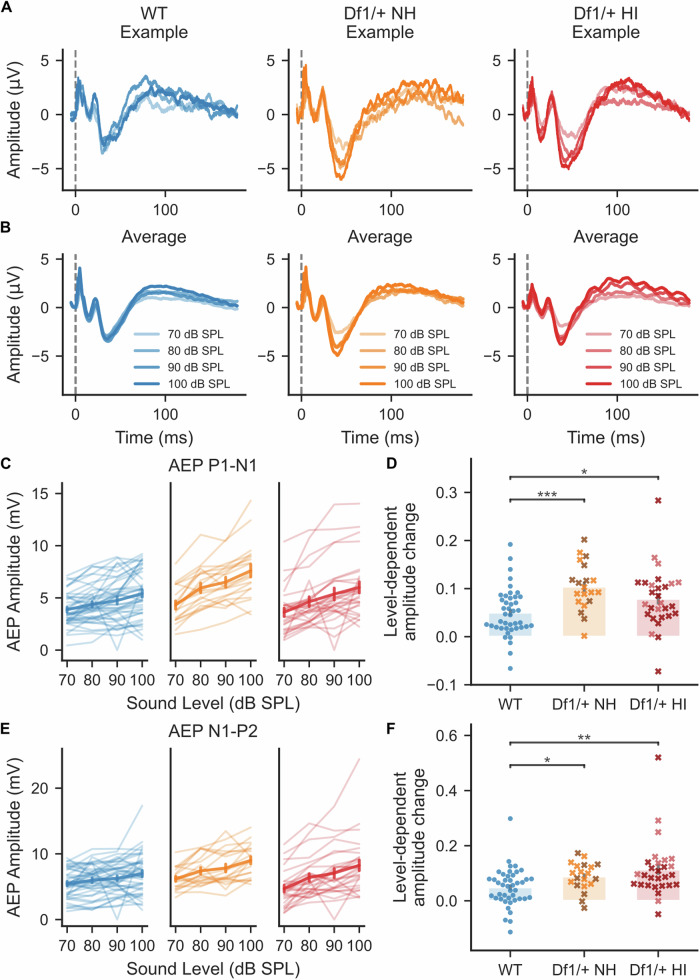
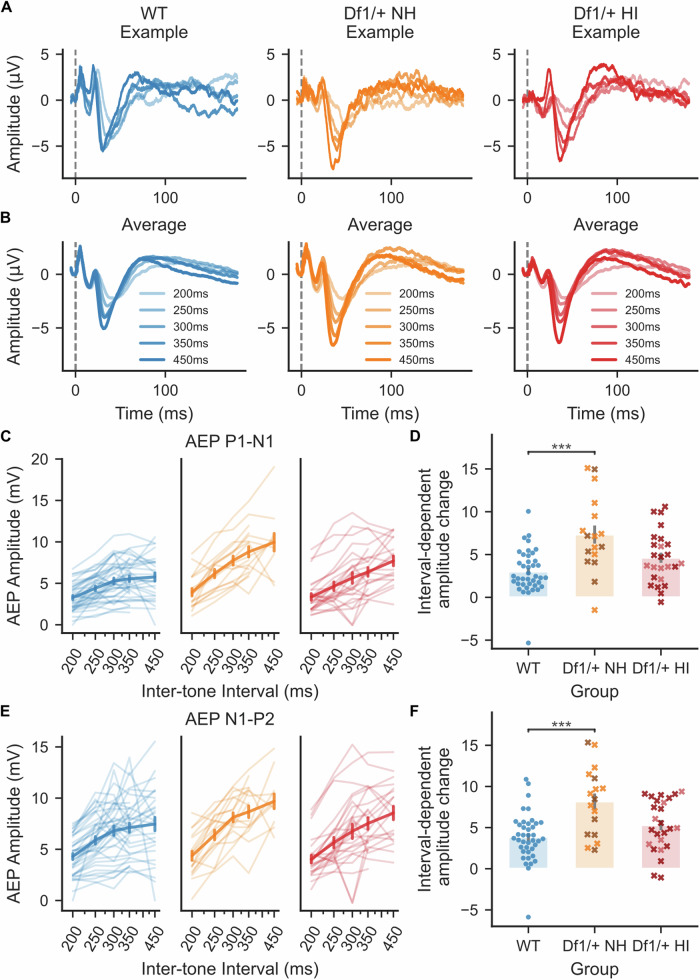
The 22q11.2 deletion is a risk factor for multiple psychiatric disorders including schizophrenia and also increases vulnerability to middle-ear problems that can cause hearing impairment. Up to 60% of deletion carriers experience hearing impairment and ~30% develop schizophrenia in adulthood. It is not known if these risks interact. Here we used the Df1/+ mouse model of the 22q11.2 deletion to investigate how hearing impairment might interact with increased genetic vulnerability to psychiatric disease to affect brain function. We measured brain function using cortical auditory evoked potentials (AEPs), which are commonly measured non-invasively in humans. After identifying one of the simplest and best-validated methods for AEP measurement in mice from the diversity of previous approaches, we measured peripheral hearing sensitivity and cortical AEPs in Df1/+ mice and their WT littermates. We exploited large inter-individual variation in hearing ability among Df1/+ mice to distinguish effects of genetic background from effects of hearing impairment. Central auditory gain and adaptation were quantified by comparing brainstem activity and cortical AEPs and by analyzing the growth of cortical AEPs with increasing sound level or inter-tone interval duration. We found that level-dependent AEP growth was abnormally large in Df1/+ mice regardless of hearing impairment, but other AEP measures of central auditory gain and adaptation depended on both genotype and hearing phenotype. Our results demonstrate the relevance of comorbid hearing loss to auditory brain dysfunction in 22q11.2DS and also identify potential biomarkers for psychiatric disease that are robust to hearing impairment.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethics approval and consent to participate: This study involved only animal research. All methods and experiments were performed in accordance with relevant guidelines and regulations of the UCL Animal Welfare and Ethical Review Body and a UK Home Office project licence approved under the United Kingdom Animals (Scientific Procedures) Act of 1986.
Figures

References
-
- Schneider M, Debbané M, Bassett AS, Chow EWC, Fung WLA, van den Bree M, et al. Psychiatric disorders from childhood to adulthood in 22q11.2 deletion syndrome: results from the International Consortium on Brain and Behavior in 22q11.2 Deletion Syndrome. Am J Psychiatry. 2014;171:627–39. - PMC - PubMed
-
- Monks S, Niarchou M, Davies AR, Walters JTR, Williams N, Owen MJ, et al. Further evidence for high rates of schizophrenia in 22q11.2 deletion syndrome. Schizophr Res. 2014;153:231–6. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
