

Endothelial tip-cell position, filopodia formation and biomechanics require BMPR2 expression and signaling
- PMID: 39779836
- PMCID: PMC11711618
- DOI: 10.1038/s42003-024-07431-8
Endothelial tip-cell position, filopodia formation and biomechanics require BMPR2 expression and signaling
Abstract
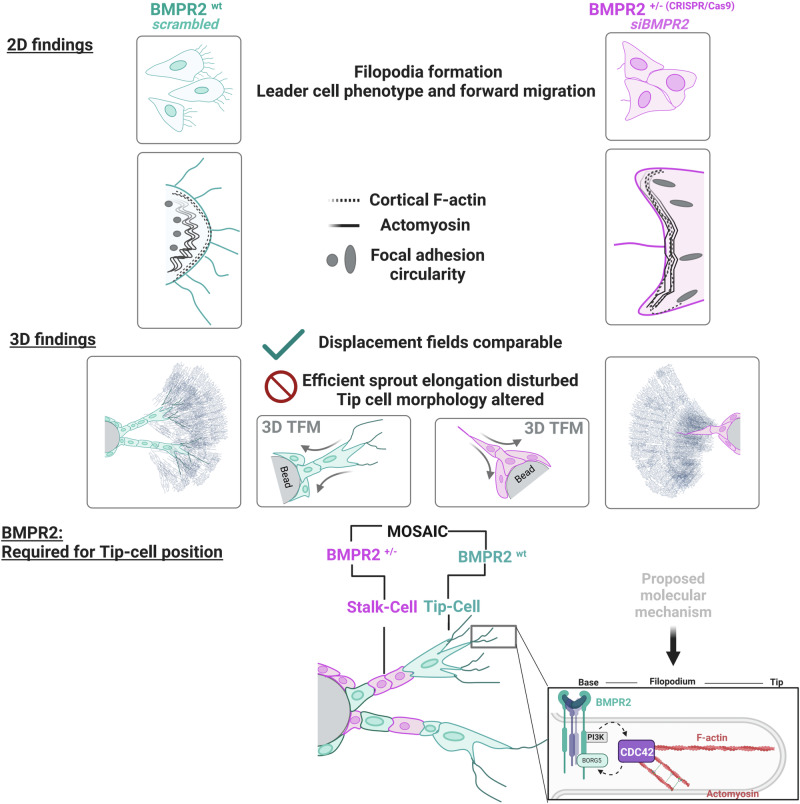
Blood vessel formation relies on biochemical and mechanical signals, particularly during sprouting angiogenesis when endothelial tip cells (TCs) guide sprouting through filopodia formation. The contribution of BMP receptors in defining tip-cell characteristics is poorly understood. Our study combines genetic, biochemical, and molecular methods together with 3D traction force microscopy, which reveals an essential role of BMPR2 for actin-driven filopodia formation and mechanical properties of endothelial cells (ECs). Targeting of Bmpr2 reduced sprouting angiogenesis in zebrafish and BMPR2-deficient human ECs formed fewer filopodia, affecting cell migration and actomyosin localization. Spheroid assays revealed a reduced sprouting of BMPR2-deficient ECs in fibrin gels. Even more strikingly, in mosaic spheroids, BMPR2-deficient ECs failed to acquire tip-cell positions. Yet, 3D traction force microscopy revealed that these distinct cell behaviors of BMPR2-deficient tip cells cannot be explained by differences in force-induced matrix deformations, even though these cells adopted distinct cone-shaped morphologies. Notably, BMPR2 positively regulates local CDC42 activity at the plasma membrane to promote filopodia formation. Our findings reveal that BMPR2 functions as a nexus integrating biochemical and biomechanical processes crucial for TCs during angiogenesis.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
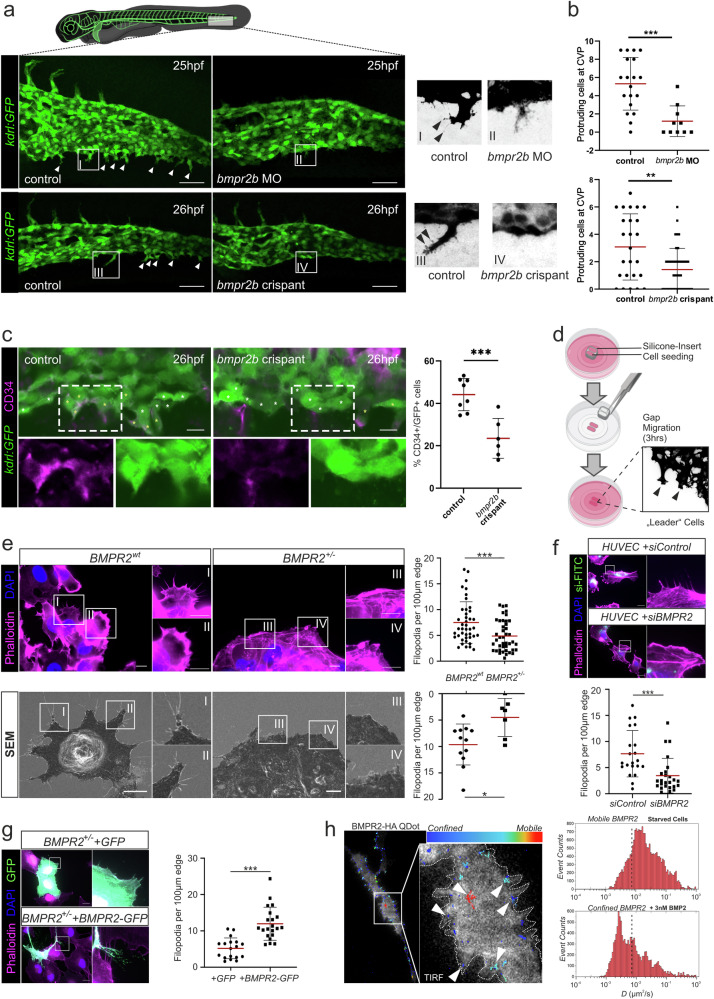
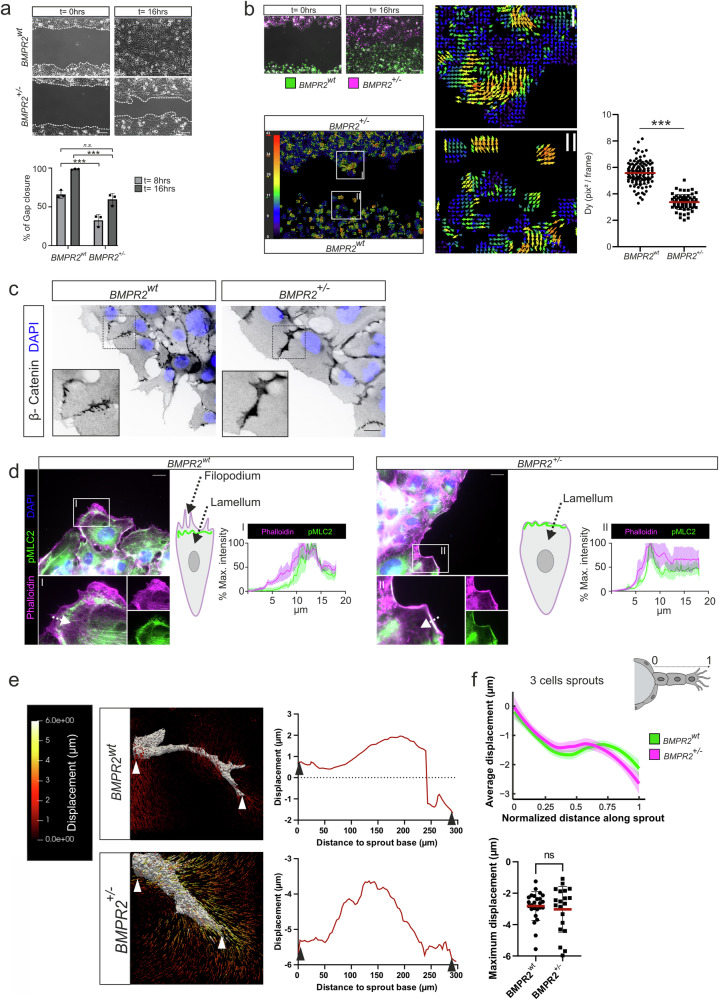
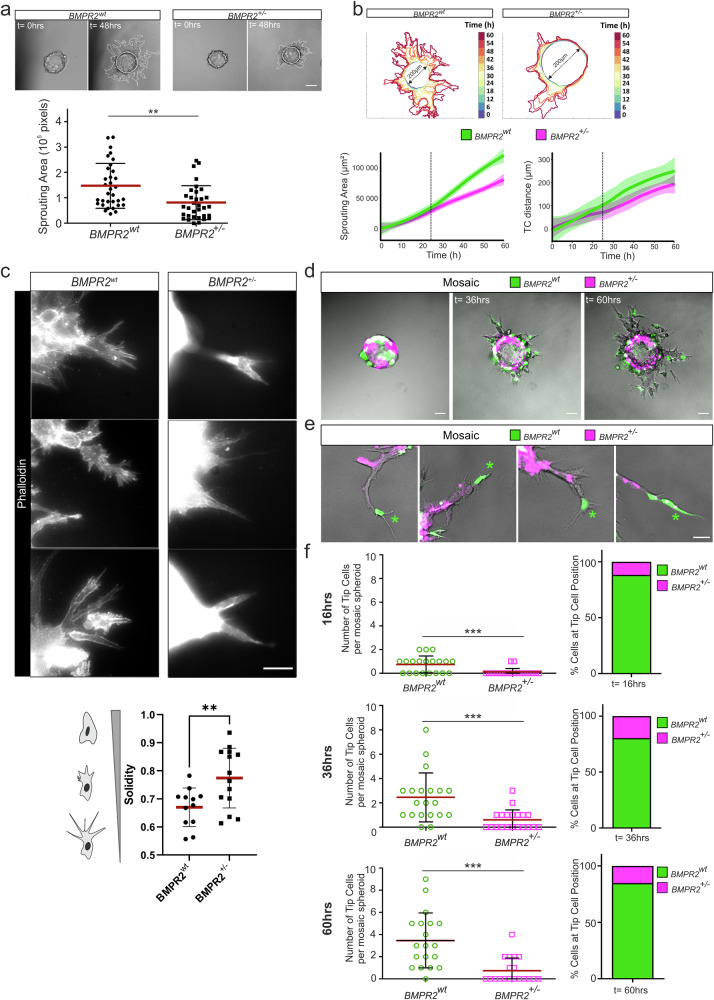
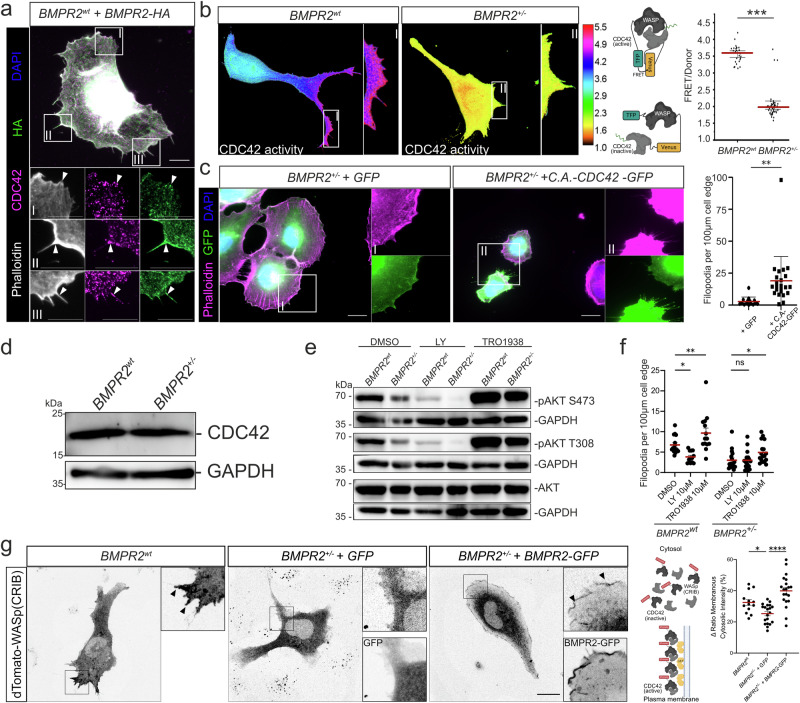
References
MeSH terms
Substances
Grants and funding
- IMPRS-BAC/Max-Planck-Gesellschaft (Max Planck Society)
- IMPRS-BAC/Max-Planck-Gesellschaft (Max Planck Society)
- BSIO/Deutsche Forschungsgemeinschaft (German Research Foundation)
- BL1514/1/Deutsche Forschungsgemeinschaft (German Research Foundation)
- BL1514/1/Deutsche Forschungsgemeinschaft (German Research Foundation)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
