

Bidirectional histone monoaminylation dynamics regulate neural rhythmicity
- PMID: 39779849
- PMCID: PMC11754111
- DOI: 10.1038/s41586-024-08371-3
Bidirectional histone monoaminylation dynamics regulate neural rhythmicity
Abstract
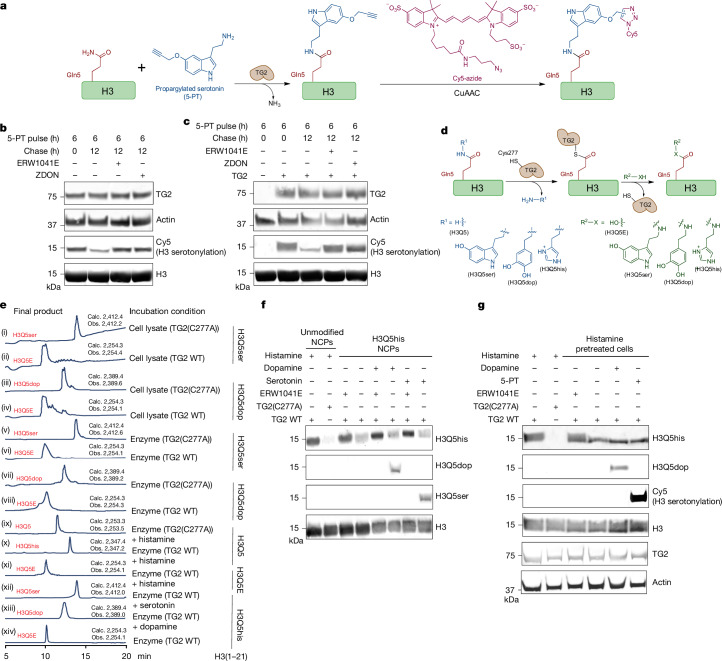
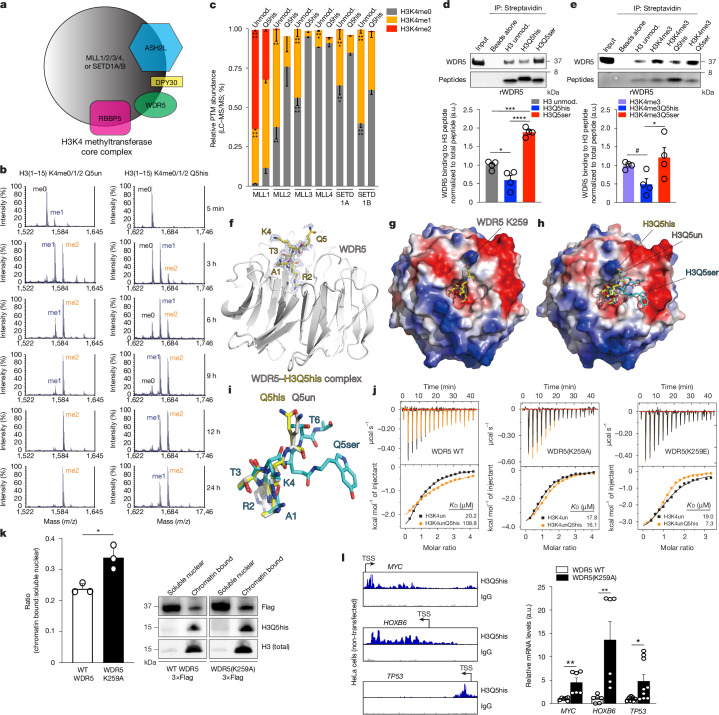
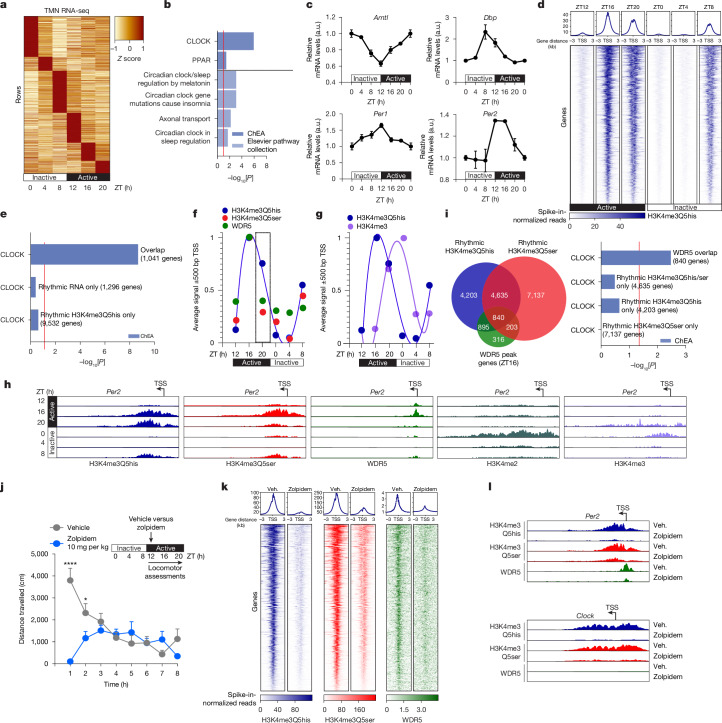
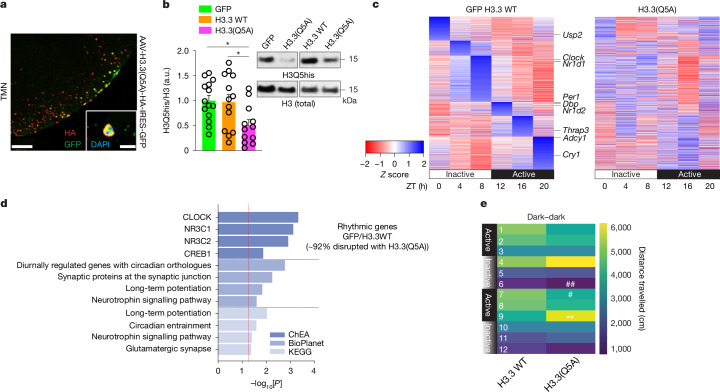
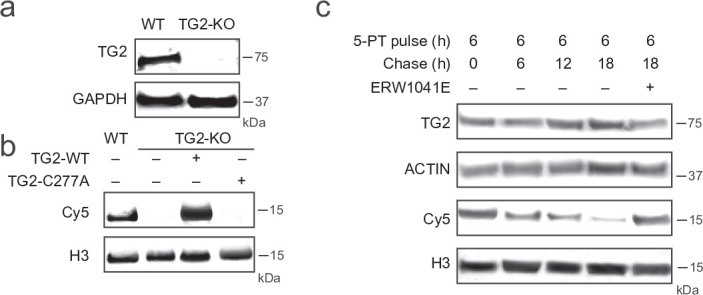
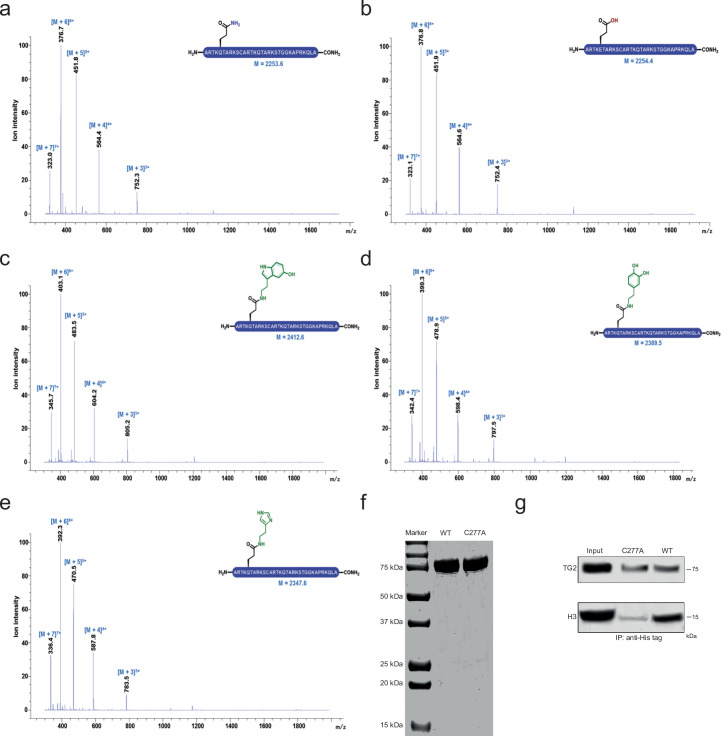
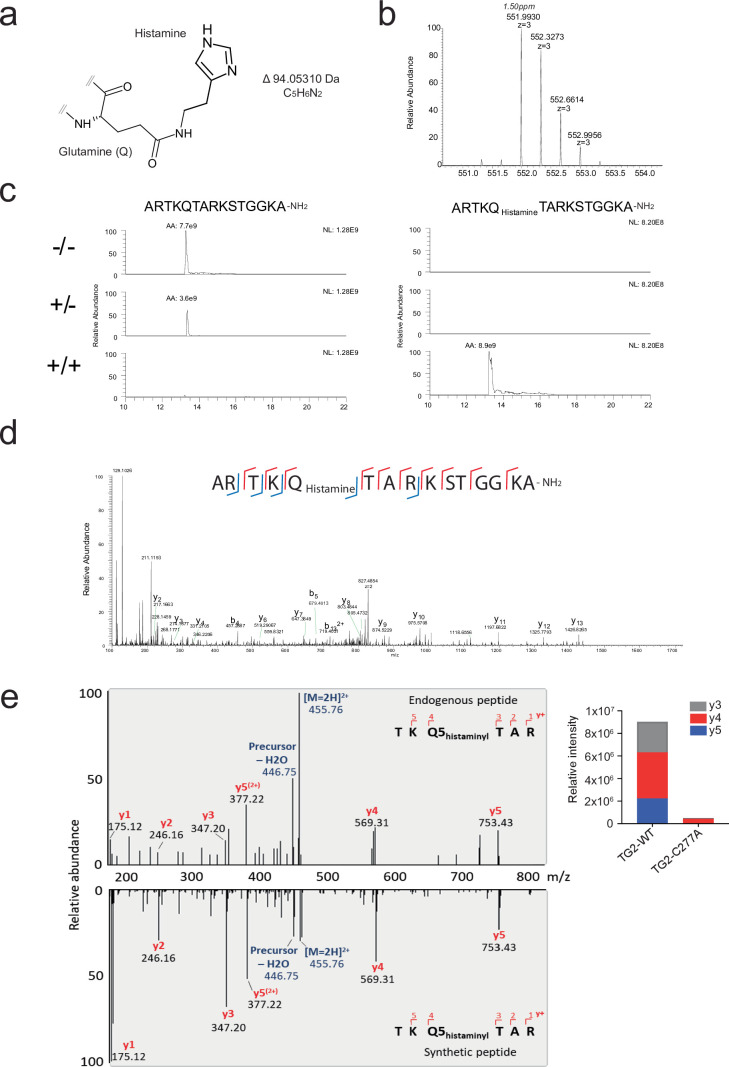
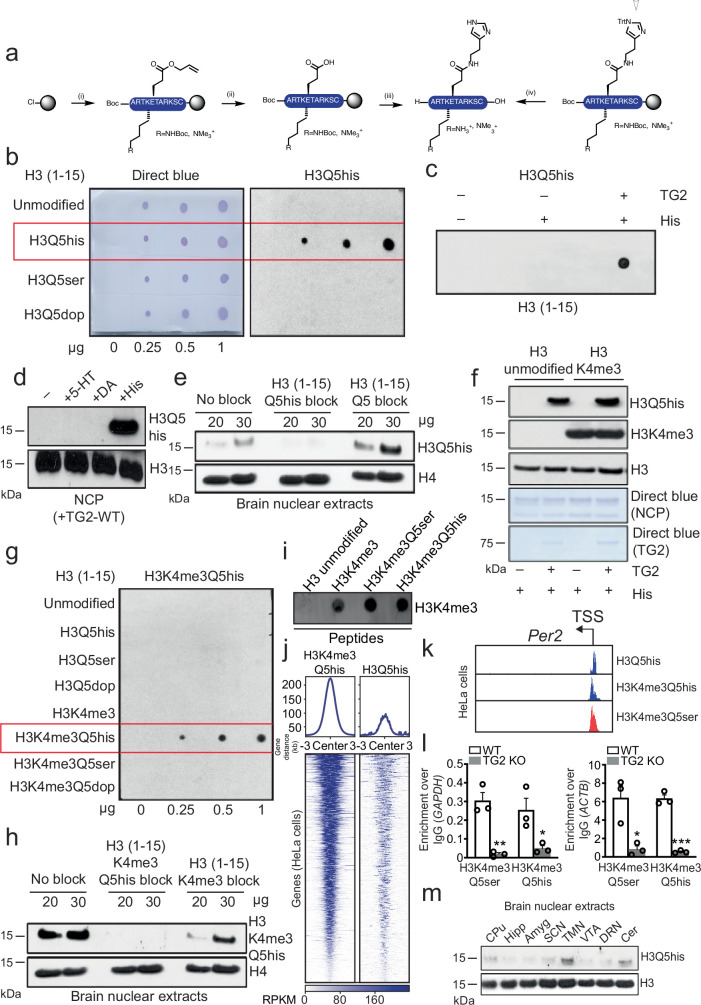
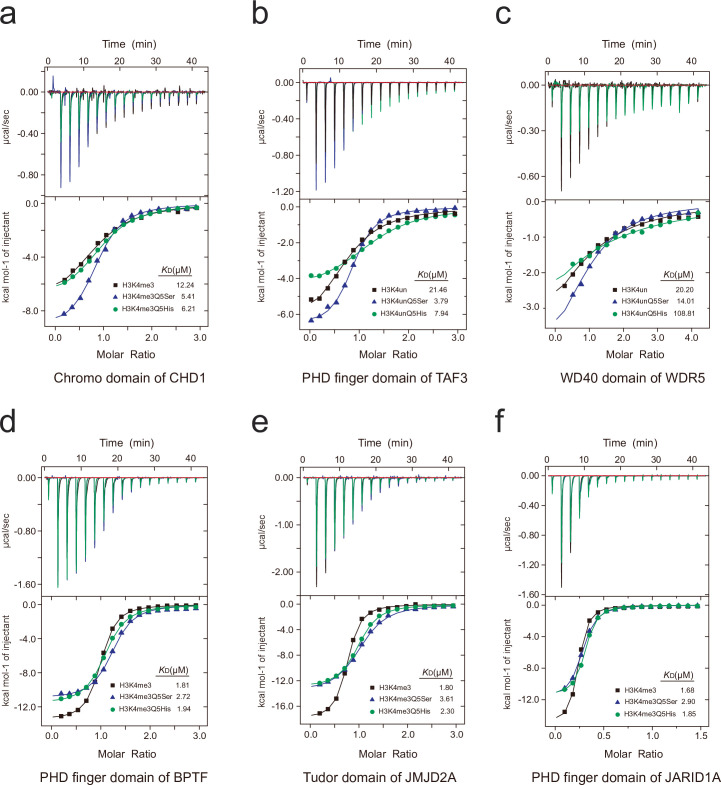
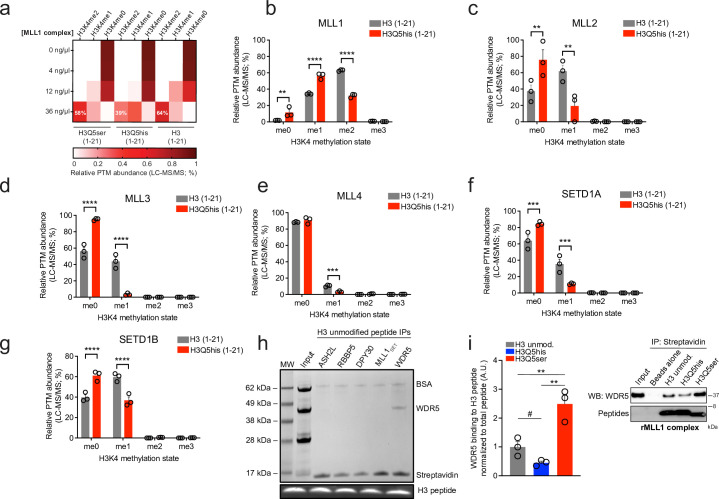
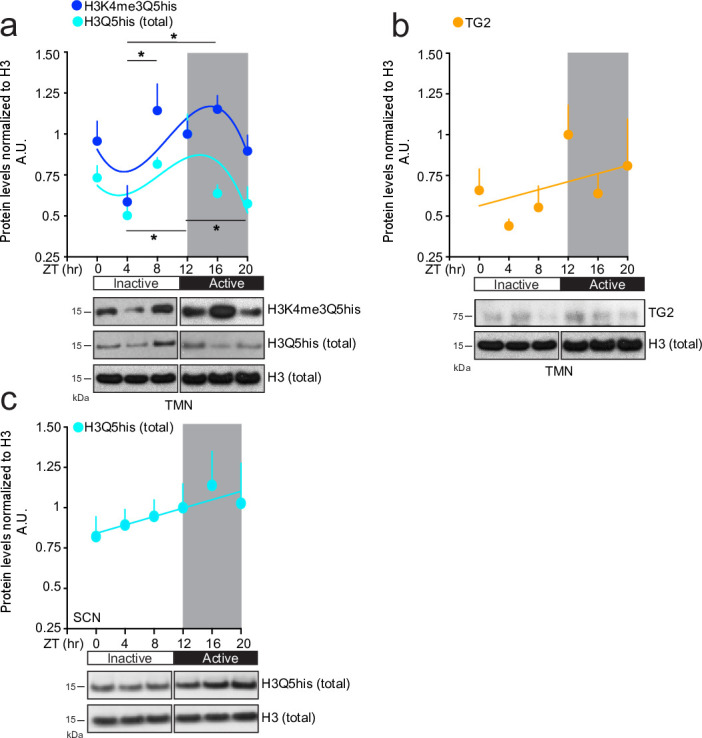
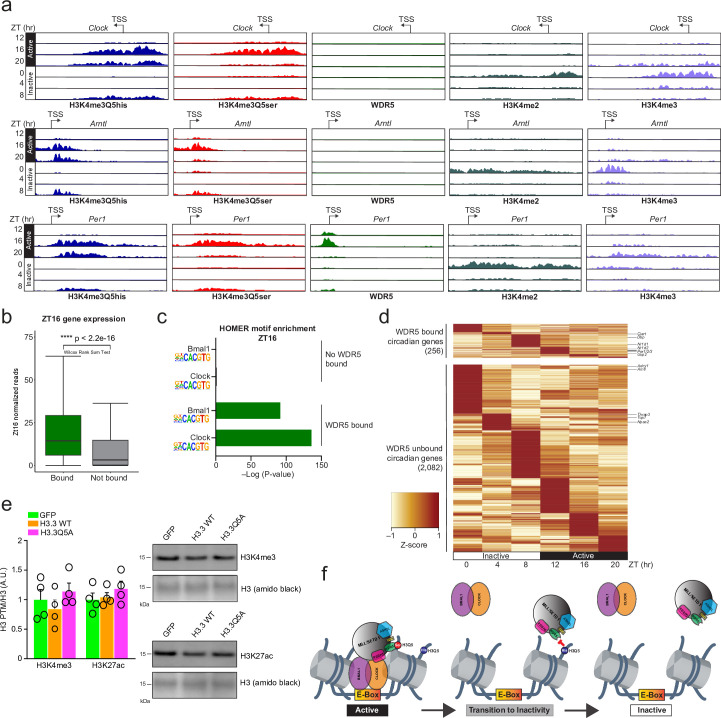
Histone H3 monoaminylations at Gln5 represent an important family of epigenetic marks in brain that have critical roles in permissive gene expression1-3. We previously demonstrated that serotonylation4-10 and dopaminylation9,11-13 of Gln5 of histone H3 (H3Q5ser and H3Q5dop, respectively) are catalysed by transglutaminase 2 (TG2), and alter both local and global chromatin states. Here we found that TG2 additionally functions as an eraser and exchanger of H3 monoaminylations, including H3Q5 histaminylation (H3Q5his), which displays diurnally rhythmic expression in brain and contributes to circadian gene expression and behaviour. We found that H3Q5his, in contrast to H3Q5ser, inhibits the binding of WDR5, a core member of histone H3 Lys4 (H3K4) methyltransferase complexes, thereby antagonizing methyltransferase activities on H3K4. Taken together, these data elucidate a mechanism through which a single chromatin regulatory enzyme has the ability to sense chemical microenvironments to affect the epigenetic states of cells, the dynamics of which have critical roles in the regulation of neural rhythmicity.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

Similar articles
-
Chemical Proteomic Profiling of Protein Dopaminylation in Colorectal Cancer Cells.J Proteome Res. 2024 Jul 5;23(7):2651-2660. doi: 10.1021/acs.jproteome.4c00379. Epub 2024 Jun 5. J Proteome Res. 2024. PMID: 38838187
-
TGM2-mediated histone serotonylation promotes HCC progression via MYC signalling pathway.J Hepatol. 2025 Jul;83(1):105-118. doi: 10.1016/j.jhep.2024.12.038. Epub 2025 Jan 7. J Hepatol. 2025. PMID: 39788430
-
Bath: a Bayesian approach to analyze epigenetic transitions reveals a dual role of H3K27me3 in chondrogenesis.Epigenetics Chromatin. 2025 Jun 27;18(1):38. doi: 10.1186/s13072-025-00594-6. Epigenetics Chromatin. 2025. PMID: 40571950 Free PMC article.
-
[Epigenetics' implication in autism spectrum disorders: A review].Encephale. 2017 Aug;43(4):374-381. doi: 10.1016/j.encep.2016.07.007. Epub 2016 Sep 28. Encephale. 2017. PMID: 27692350 French.
-
Stage-specific DNA methylation dynamics in mammalian heart development.Epigenomics. 2025 Apr;17(5):359-371. doi: 10.1080/17501911.2025.2467024. Epub 2025 Feb 21. Epigenomics. 2025. PMID: 39980349 Review.
Cited by
-
Persistent dopamine-dependent remodeling of the neural transcriptome in response to pregnancy and postpartum.bioRxiv [Preprint]. 2025 Jun 2:2025.02.20.639313. doi: 10.1101/2025.02.20.639313. bioRxiv. 2025. PMID: 40060435 Free PMC article. Preprint.
-
Molecular determinants for recognition of serotonylated chromatin.Nucleic Acids Res. 2025 Jul 8;53(13):gkaf612. doi: 10.1093/nar/gkaf612. Nucleic Acids Res. 2025. PMID: 40637225 Free PMC article.
-
Phenethylaminylation: Preliminary In Vitro Evidence for the Covalent Transamidation of Psychedelic Phenethylamines to Glial Proteins using 3,5-Dimethoxy-4-(2-Propynyloxy)-Phenethylamine as a Model Compound.bioRxiv [Preprint]. 2025 Feb 17:2025.02.13.638188. doi: 10.1101/2025.02.13.638188. bioRxiv. 2025. PMID: 40027829 Free PMC article. Preprint.
-
Research progress on cancer-related epigenetic switches.Invest New Drugs. 2025 Jun;43(3):525-559. doi: 10.1007/s10637-025-01555-2. Epub 2025 Jun 13. Invest New Drugs. 2025. PMID: 40512440 Review.
-
Macrophage clock of pregnancy: circadian and metabolic control of decidual macrophage.Semin Immunopathol. 2025 Jul 18;47(1):30. doi: 10.1007/s00281-025-01057-6. Semin Immunopathol. 2025. PMID: 40679680 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
