

Structural polymorphism and diversity of human segmental duplications
- PMID: 39779957
- PMCID: PMC11821543
- DOI: 10.1038/s41588-024-02051-8
Structural polymorphism and diversity of human segmental duplications
Abstract
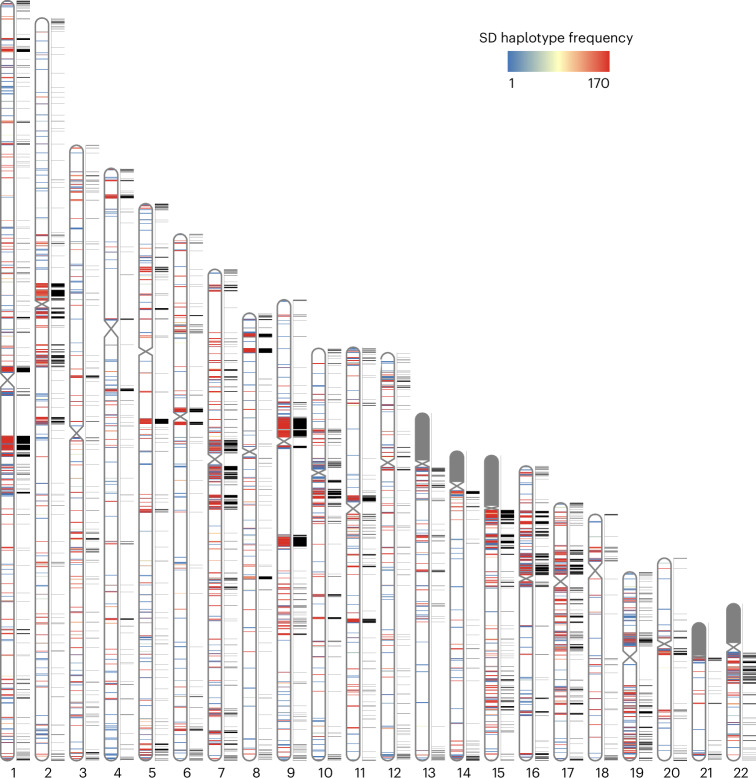
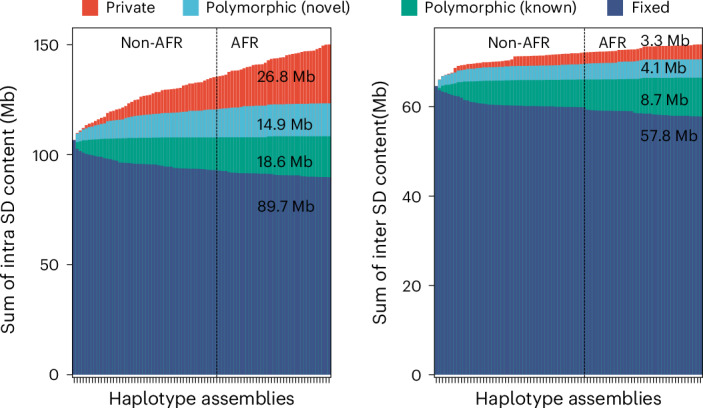
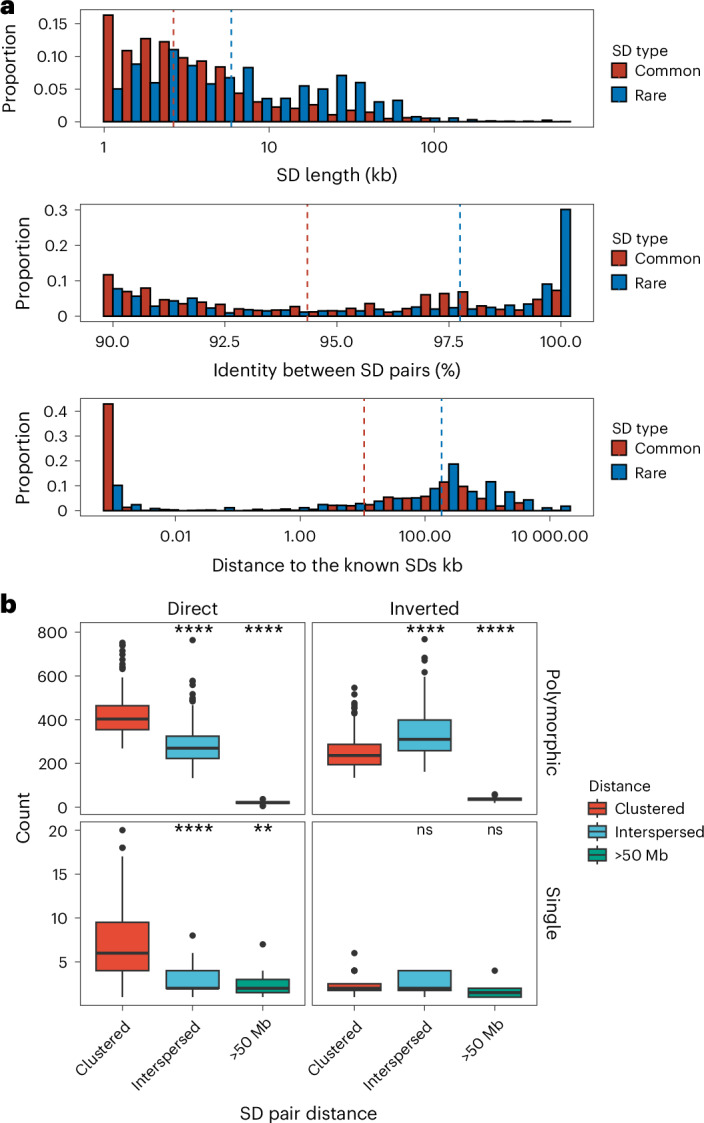
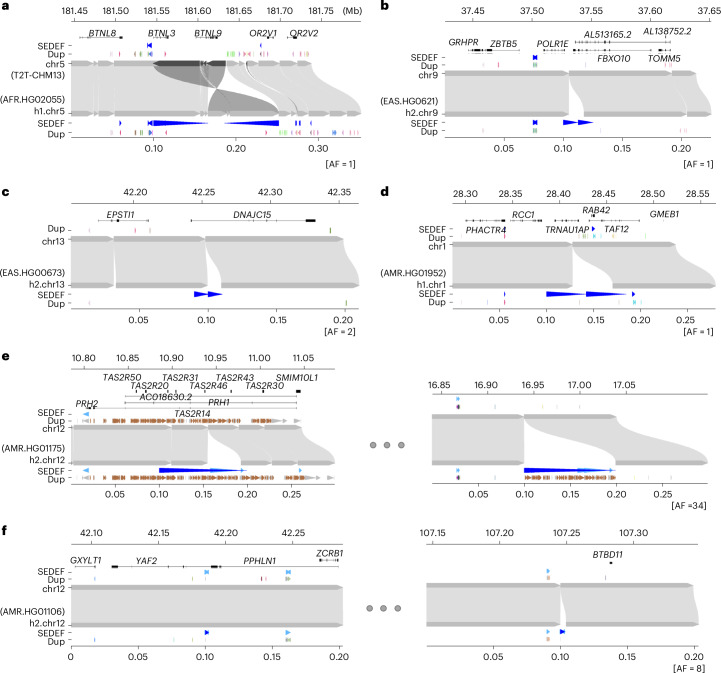
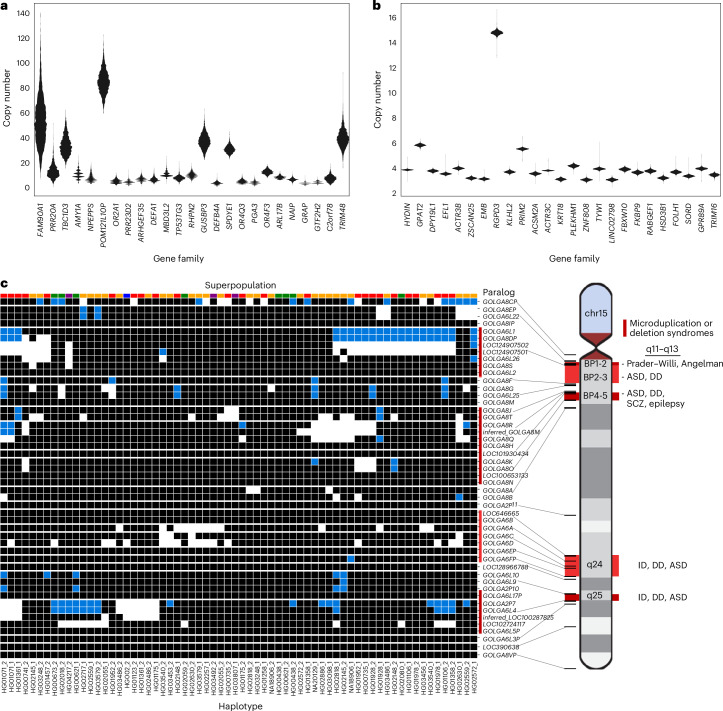
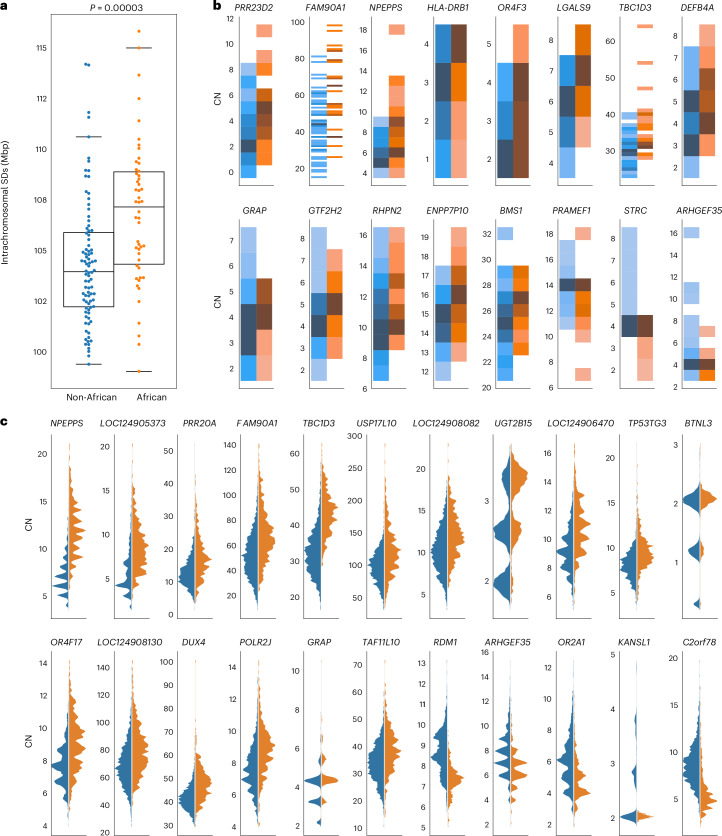
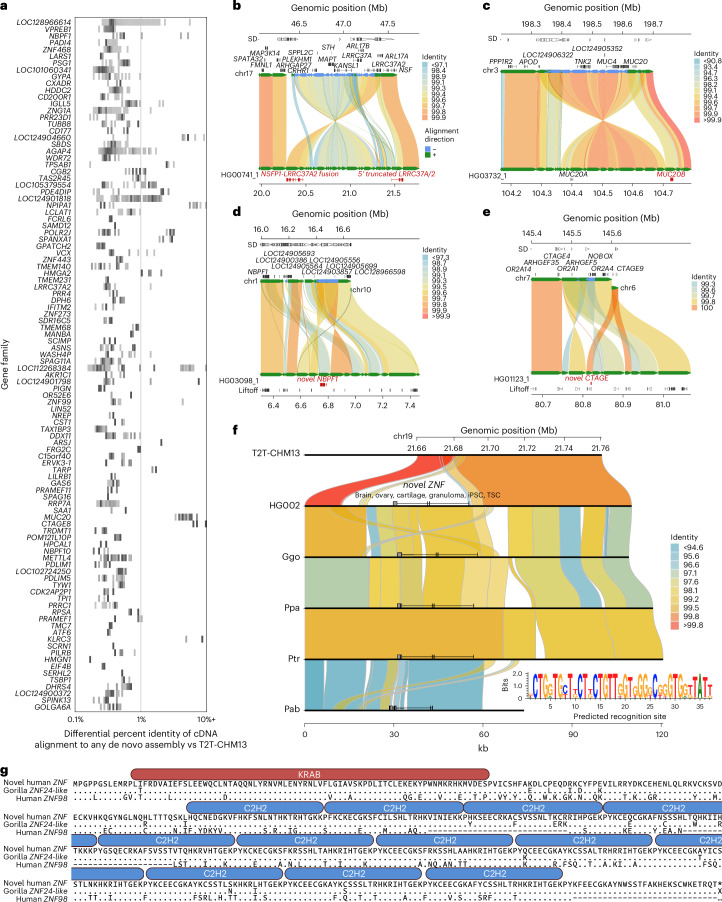
Segmental duplications (SDs) contribute significantly to human disease, evolution and diversity but have been difficult to resolve at the sequence level. We present a population genetics survey of SDs by analyzing 170 human genome assemblies (from 85 samples representing 38 Africans and 47 non-Africans) in which the majority of autosomal SDs are fully resolved using long-read sequence assembly. Excluding the acrocentric short arms and sex chromosomes, we identify 173.2 Mb of duplicated sequence (47.4 Mb not present in the telomere-to-telomere reference) distinguishing fixed from structurally polymorphic events. We find that intrachromosomal SDs are among the most variable, with rare events mapping near their progenitor sequences. African genomes harbor significantly more intrachromosomal SDs and are more likely to have recently duplicated gene families with higher copy numbers than non-African samples. Comparison to a resource of 563 million full-length isoform sequencing reads identifies 201 novel, potentially protein-coding genes corresponding to these copy number polymorphic SDs.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: E.E.E. is a scientific advisory board member of Variant Bio. C.L. is a scientific advisory board member of Nabsys and Genome Insight. The other authors declare no competing interests.
Figures
Update of
-
Structural polymorphism and diversity of human segmental duplications.bioRxiv [Preprint]. 2024 Jun 6:2024.06.04.597452. doi: 10.1101/2024.06.04.597452. bioRxiv. 2024. Update in: Nat Genet. 2025 Feb;57(2):390-401. doi: 10.1038/s41588-024-02051-8. PMID: 38895457 Free PMC article. Updated. Preprint.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
