

PRDX5 and PRDX6 translocation and oligomerization in bull sperm: a response to cryopreservation-induced oxidative stress
- PMID: 39780184
- PMCID: PMC11714857
- DOI: 10.1186/s12964-024-02015-9
PRDX5 and PRDX6 translocation and oligomerization in bull sperm: a response to cryopreservation-induced oxidative stress
Abstract
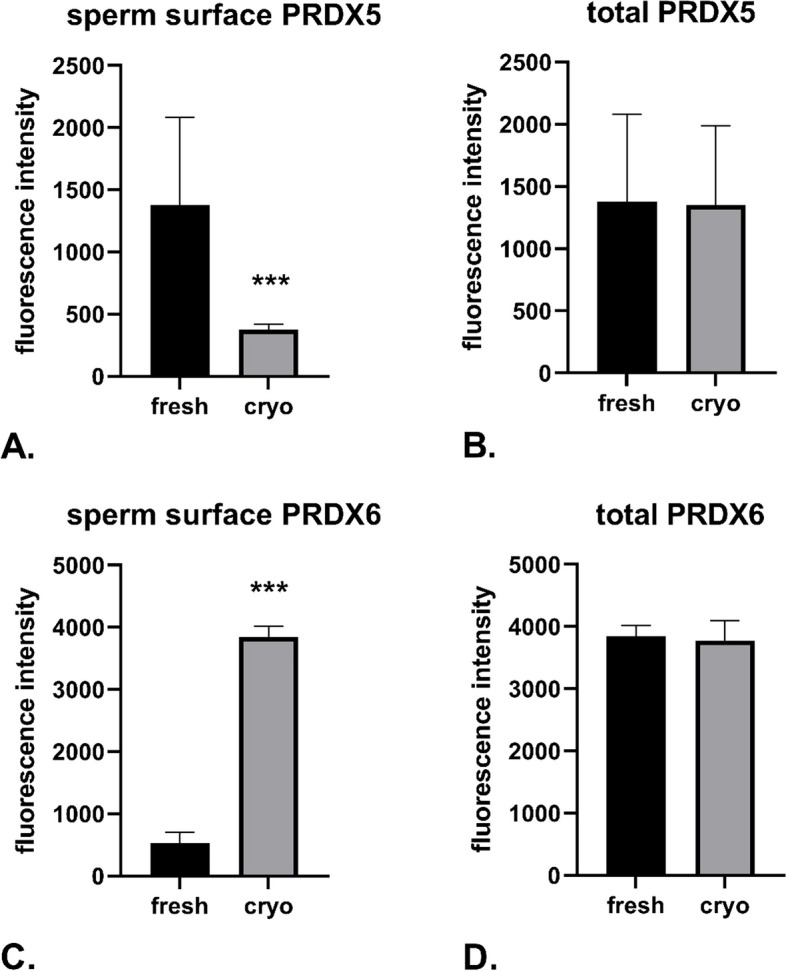
Cryopreservation of bull sperm, crucial for breeding and assisted reproduction, often reduces sperm quality due to oxidative stress. This study examines how oxidative stress during cryopreservation affects peroxiredoxin 5 (PRDX5) and peroxiredoxin 6 (PRDX6) proteins, leading to their translocation and oligomerization in bull sperm. Increased reactive oxygen species (ROS) and nitric oxide (NO) levels were linked to reduced mitochondrial potential, higher DNA fragmentation, and increased membrane fluidity, prompting PRDX5 to move intracellularly and PRDX6 to the cell membrane. Under cryopreservation, these proteins formed high molecular weight oligomers, that may shift from peroxidase to chaperone roles. Their interaction with Toll-like receptor 4 (TLR4) may be key to their intracellular transport. On the other hand, the presence of PRDX5 and PRDX6 in exosomal vesicles suggested a potential mechanism for their transport into sperm cells. Using Imaging Flow Cytometry and various PAGE techniques, the study detected PRDX5 and PRDX6 in different sperm locations and analyzed their oligomer formation. These findings highlight the adaptive roles of PRDX5 and PRDX6 in protecting sperm cells, offering insights that could improve cryopreservation protocols in animal breeding and human reproductive medicine, and advance our understanding of the oxidative stress response in sperm cells.
Keywords: Bull sperm; Cryopreservation; Oxidative stress; PRDX5; PRDX6.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures







References
-
- Mostek A, Dietrich MA, Słowińska M, Ciereszko A. Cryopreservation of bull semen is associated with carbonylation of sperm proteins. Theriogenology. 2017;92:95–102. Available from: http://linkinghub.elsevier.com/retrieve/pii/S0093691X17300110 - PubMed
-
- Xin L, Guo Y, Zhao HB, Yu HM, Hou L. Peroxiredoxin 6 translocates to the plasma membrane of human sperm under oxidative stress during cryopreservation. Cryobiology. 2021;100:158–63. - PubMed
-
- Rezaei N, Mohammadi M, Mohammadi H, Khalatbari A, Zare Z. Acrosome and chromatin integrity, oxidative stress, and expression of apoptosis-related genes in cryopreserved mouse epididymal spermatozoa treated with L-Carnitine. Cryobiology. 2020;95:171–6. - PubMed
-
- Aitken RJ, Gibb Z, Baker MA, Drevet J, Gharagozloo P. Causes and consequences of oxidative stress in Spermatozoa. Reprod Fertil Dev. 2016;28:1. - PubMed
-
- Knoops B, Goemaere J, Van Der Eecken V, Declercq JP. Peroxiredoxin 5: Structure, mechanism, and function of the mammalian atypical 2-cys peroxiredoxin. Antioxid Redox Signal. 2011;15:817–29. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
