

Alleviation of liver fibrosis by inhibiting a non-canonical ATF4-regulated enhancer program in hepatic stellate cells
- PMID: 39789010
- PMCID: PMC11718104
- DOI: 10.1038/s41467-024-55738-1
Alleviation of liver fibrosis by inhibiting a non-canonical ATF4-regulated enhancer program in hepatic stellate cells
Abstract
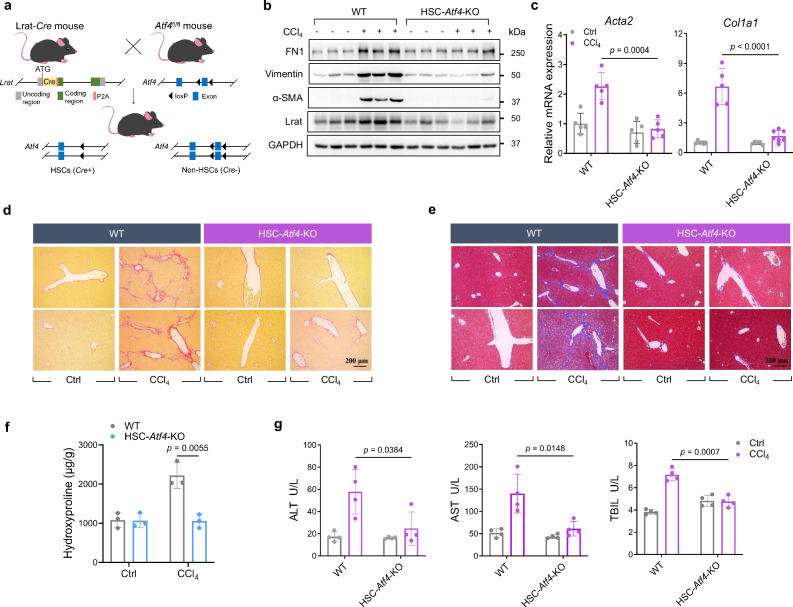
Liver fibrosis is a critical liver disease that can progress to more severe manifestations, such as cirrhosis, yet no effective targeted therapies are available. Here, we identify that ATF4, a master transcription factor in ER stress response, promotes liver fibrosis by facilitating a stress response-independent epigenetic program in hepatic stellate cells (HSCs). Unlike its canonical role in regulating UPR genes during ER stress, ATF4 activates epithelial-mesenchymal transition (EMT) gene transcription under fibrogenic conditions. HSC-specific depletion of ATF4 suppresses liver fibrosis in vivo. Mechanistically, TGFβ resets ATF4 to orchestrate a unique enhancer program for the transcriptional activation of pro-fibrotic EMT genes. Analysis of human data confirms a strong correlation between HSC ATF4 expression and liver fibrosis progression. Importantly, a small molecule inhibitor targeting ATF4 translation effectively mitigates liver fibrosis. Together, our findings identify a mechanism promoting liver fibrosis and reveal new opportunities for treating this otherwise non-targetable disease.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures






References
-
- Hernandez-Gea, V. & Friedman, S. L. Pathogenesis of liver fibrosis. Annu Rev. Pathol.6, 425–456 (2011). - PubMed
-
- Kisseleva, T. & Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat. Rev. Gastroenterol. Hepatol.18, 151–166 (2021). - PubMed
-
- Cotter, T. G. & Rinella, M. Fatty liver disease 2020: The state of the disease. Gastroenterology158, 1851–1864 (2020). - PubMed
-
- Parola, M. & Pinzani, M. Liver fibrosis: Pathophysiology, pathogenetic targets and clinical issues. Mol. Asp. Med.65, 37–55 (2019). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
