

The origin of color categories
- PMID: 39793082
- PMCID: PMC11725794
- DOI: 10.1073/pnas.2400273121
The origin of color categories
Abstract
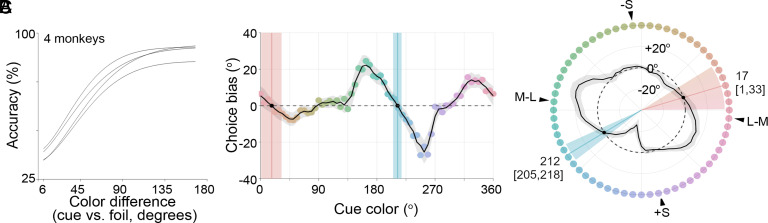
To what extent does concept formation require language? Here, we exploit color to address this question and ask whether macaque monkeys have color concepts evident as categories. Macaques have similar cone photoreceptors and central visual circuits to humans, yet they lack language. Whether Old World monkeys such as macaques have consensus color categories is unresolved, but if they do, then language cannot be required. If macaques do not have color categories, then color categories in humans are unlikely to derive from innate properties of visual encoding and likely to depend on cognitive abilities such as language that differ between monkeys and humans. We tested macaques by adapting a match-to-sample paradigm used in humans to uncover color categories from errors in matches, and we analyzed the data using computational simulations that assess the possibility of unrecognized distortions in the perceptual uniformity of color space. The results provide evidence that humans have consensus cognitive color categories and macaques do not. One animal showed evidence for a private color category, demonstrating that monkeys have the capacity to form color categories even if they do not form consensus color categories. Taken together, the results imply that consensus color categories in humans, for which there is ample evidence, must depend upon language or other cognitive abilities.
Keywords: categorization; color; language.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures






References
-
- Berlin B., Kay P., Basic Color Terms: Their Universality and Evolution (University of California Press, Berkeley, CA, 1969).
-
- Heider E. R., Universals in color naming and memory. J. Exp. Psychol. 93, 10–20 (1972). - PubMed
-
- Bornstein M. H., Kessen W., Weiskopf S., The Categories of Hue in Infancy. Science 191, 201–202 (1976). - PubMed
-
- Kay P., McDaniel C. K., The linguistic significance of the meanings of basic color terms. Language 54, 610–646 (1978).
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
