

Post-transcriptional cross- and auto-regulation buffer expression of the human RNA helicases DDX3X and DDX3Y
- PMID: 39794123
- PMCID: PMC11789639
- DOI: 10.1101/gr.279707.124
Post-transcriptional cross- and auto-regulation buffer expression of the human RNA helicases DDX3X and DDX3Y
Abstract
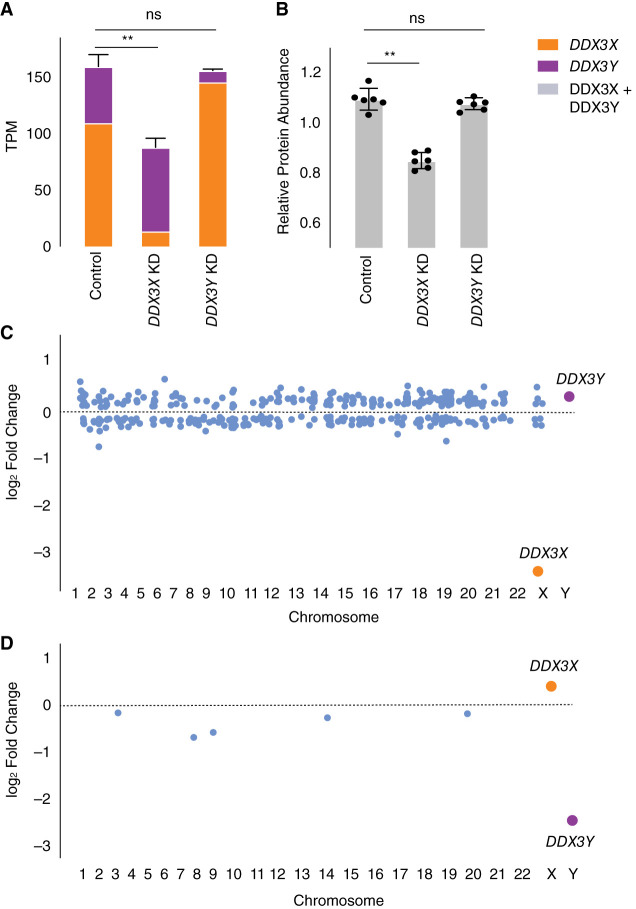
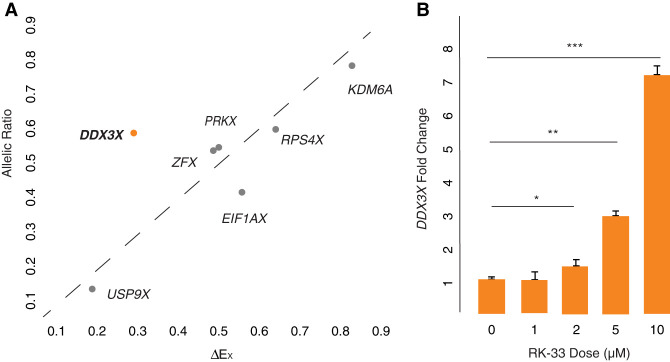
The Y-linked gene DDX3Y and its X-linked homolog DDX3X survived the evolution of the human sex chromosomes from ordinary autosomes. DDX3X encodes a multifunctional RNA helicase, with mutations causing developmental disorders and cancers. We find that, among X-linked genes with surviving Y homologs, DDX3X is extraordinarily dosage sensitive. Studying cells of individuals with sex chromosome aneuploidy, we observe that when the number of Y Chromosomes increases, DDX3X transcript levels fall; conversely, when the number of X Chromosomes increases, DDX3Y transcript levels fall. In 46,XY cells, CRISPRi knockdown of either DDX3X or DDX3Y causes transcript levels of the homologous gene to rise. In 46,XX cells, chemical inhibition of DDX3X protein activity elicits an increase in DDX3X transcript levels. Thus, perturbation of either DDX3X or DDX3Y expression is buffered: by negative cross-regulation of DDX3X and DDX3Y in 46,XY cells and by negative auto-regulation of DDX3X in 46,XX cells. DDX3X-DDX3Y cross-regulation is mediated through mRNA destabilization-as shown by metabolic labeling of newly transcribed RNA-and buffers total levels of DDX3X and DDX3Y protein in human cells. We infer that post-transcriptional auto-regulation of the ancestral (autosomal) DDX3X gene transmuted into auto- and cross-regulation of DDX3X and DDX3Y as these sex-linked genes evolved from ordinary alleles of their autosomal precursor.
© 2025 Rengarajan et al.; Published by Cold Spring Harbor Laboratory Press.
Figures





Update of
-
Post-transcriptional cross- and auto-regulation buffer expression of the human RNA helicases DDX3X and DDX3Y.bioRxiv [Preprint]. 2024 Jul 10:2024.07.08.602613. doi: 10.1101/2024.07.08.602613. bioRxiv. 2024. Update in: Genome Res. 2025 Jan 22;35(1):20-30. doi: 10.1101/gr.279707.124. PMID: 39026797 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources