

Hemolymph microbiota and immune effectors' expressions driven by geographical rearing acclimation of the aquacultured Penaeus stylirostris
- PMID: 39799372
- PMCID: PMC11725212
- DOI: 10.1186/s42523-025-00376-1
Hemolymph microbiota and immune effectors' expressions driven by geographical rearing acclimation of the aquacultured Penaeus stylirostris
Abstract
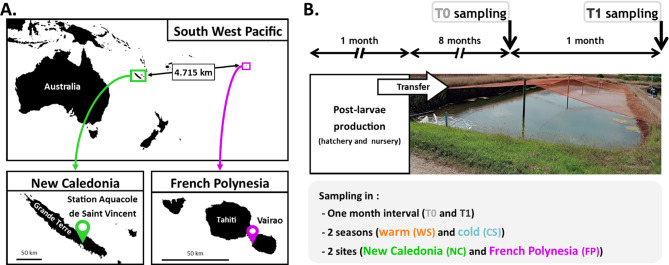
Background: In holobiont, microbiota is known to play a central role on the health and immunity of its host. Then, understanding the microbiota, its dynamic according to the environmental conditions and its link to the immunity would help to react to potential dysbiosis of aquacultured species. While the gut microbiota is highly studied, in marine invertebrates the hemolymph microbiota is often set aside even if it remains an important actor of the hemolymph homeostasis. Indeed, the hemolymph harbors the factors involved in the animal homeostasis that interacts with the microbiota, the immunity. In the Southwest Pacific, the high economical valued shrimp Penaeus stylirostris is reared in two contrasted sites, in New Caledonia (NC) and in French Polynesia (FP).
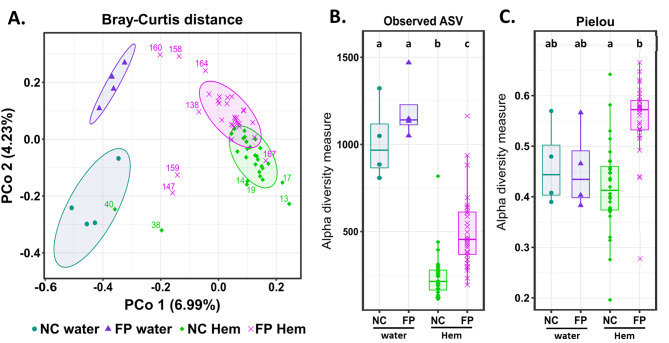
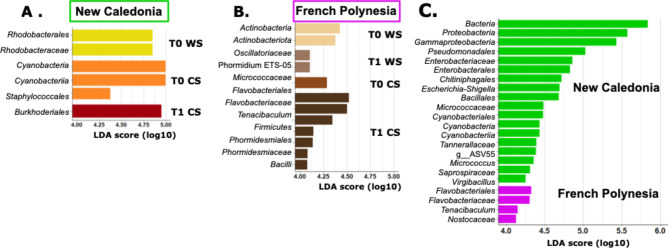
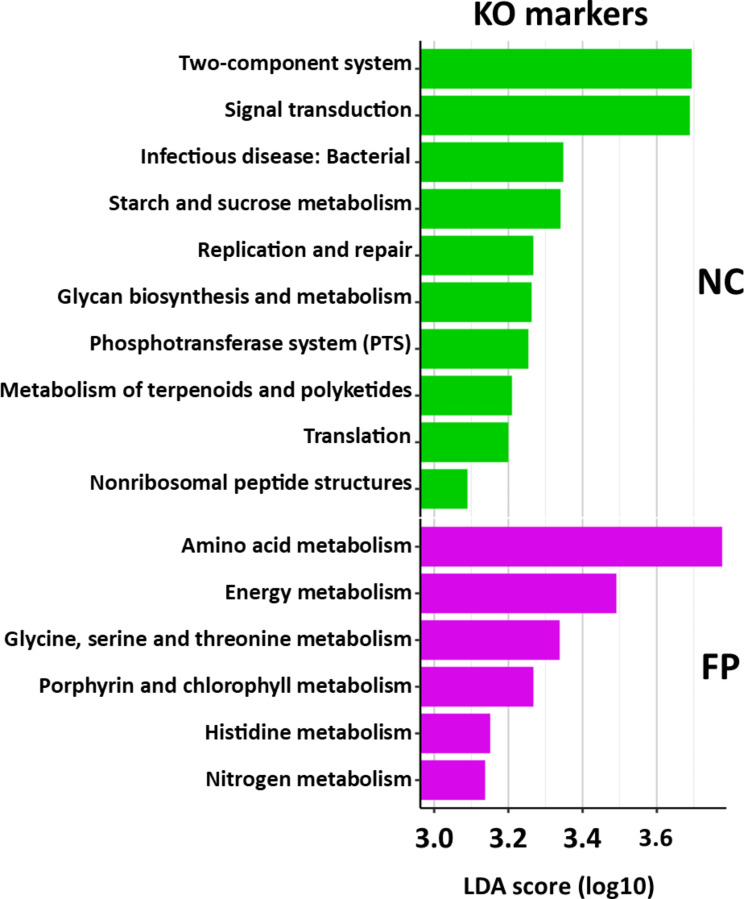
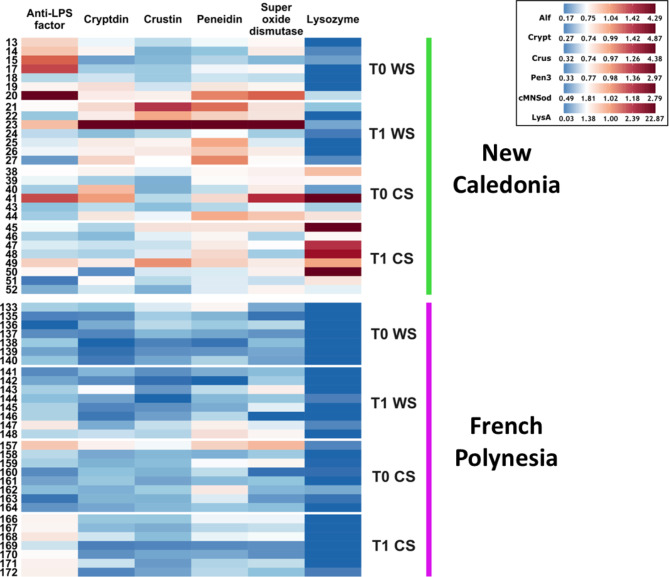
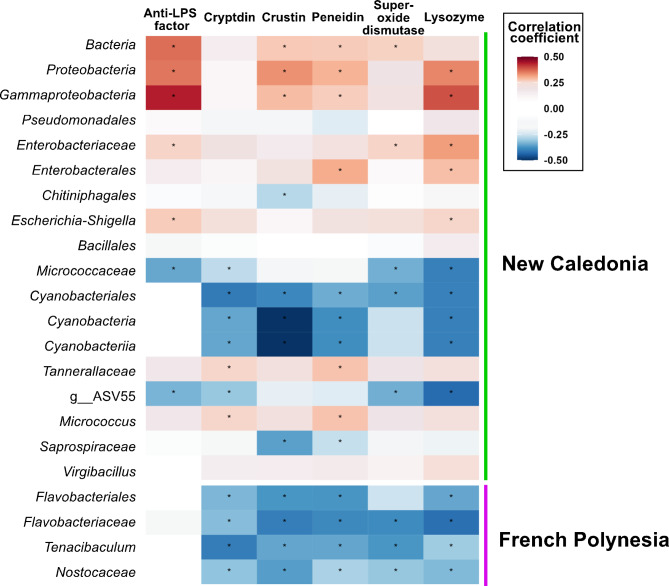
Results: We characterized the active microbiota inhabiting the hemolymph of shrimps while considering its stability during two seasons and at a one-month interval and evidenced an important microbial variability between the shrimps according to the rearing conditions and the sites. We highlighted specific biomarkers along with a common core microbiota composed of 6 ASVs. Putative microbial functions were mostly associated with bacterial competition, infections and metabolism in NC, while they were highly associated with the cell metabolism in FP suggesting a rearing site discrimination. Differential relative expression of immune effectors measured in the hemolymph of two shrimp populations from NC and FP, exhibited higher level of expression in NC compared to FP. In addition, differential relative expression of immune effectors was correlated to bacterial biomarkers based on their geographical location.
Conclusions: Our data suggest that, in Pacific shrimps, both the microbiota and the expression of the immune effectors could have undergone differential immunostimulation according to the rearing site as well as a geographical adaptative divergence of the shrimps as an holobiont, to their rearing sites. Further, the identification of proxies such as the core microbiota and site biomarkers, could be used to guide future actions to monitor the bacterial microbiota and thus preserve the productions.
Keywords: Active microbiota; Biomarkers; Geographical divergence; Host-microbiota interactions; Immune effectors; Immunostimulation; Shrimp hemolymph.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures



References
-
- Apprill A. Marine Animal microbiomes: toward understanding host–microbiome interactions in a changing ocean. Front Mar Sci. 2017;4:222. - DOI
LinkOut - more resources
Full Text Sources