

Depletion of alloreactive B cells by drug-resistant chimeric alloantigen receptor T cells to prevent transplant rejection
- PMID: 39799394
- PMCID: PMC11897811
- DOI: 10.1016/j.ymthe.2025.01.009
Depletion of alloreactive B cells by drug-resistant chimeric alloantigen receptor T cells to prevent transplant rejection
Abstract
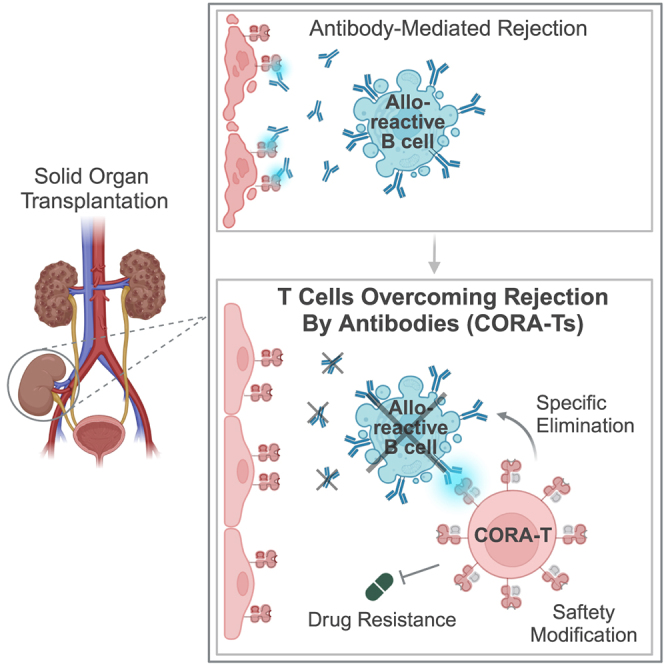
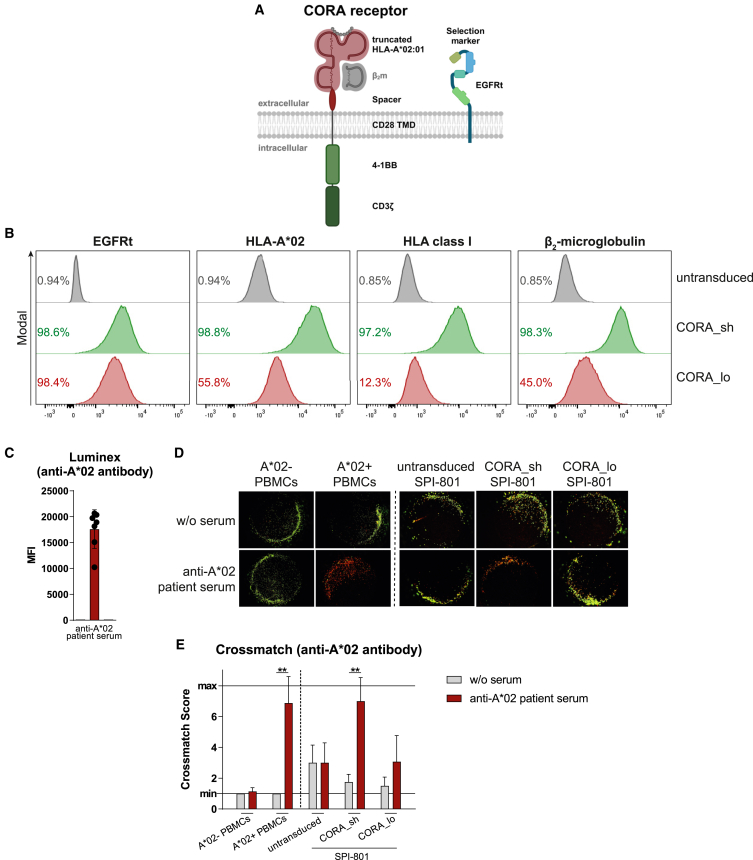
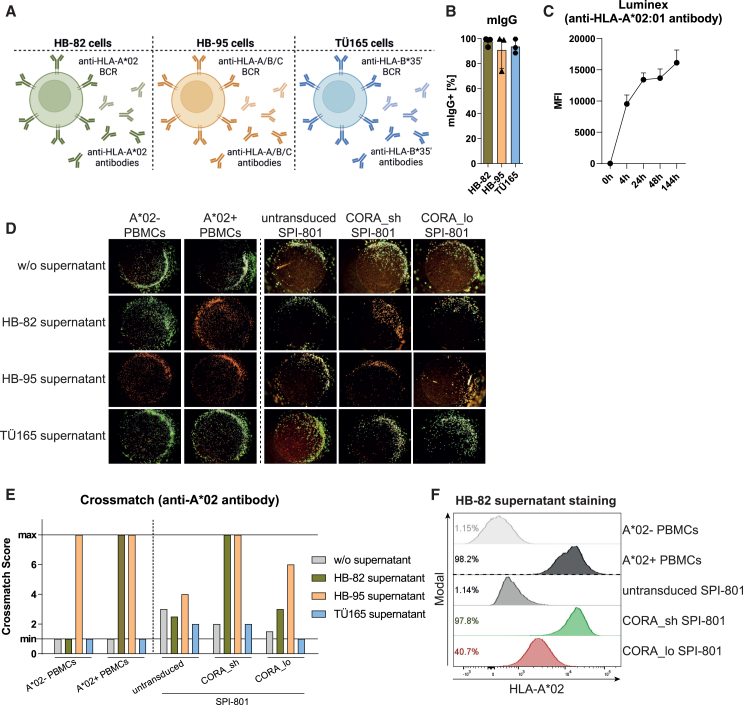
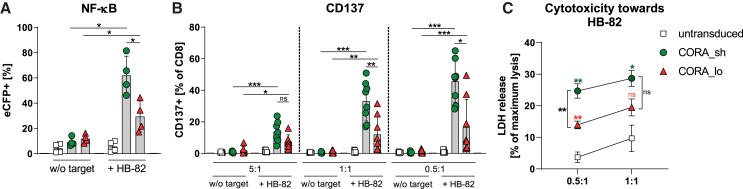
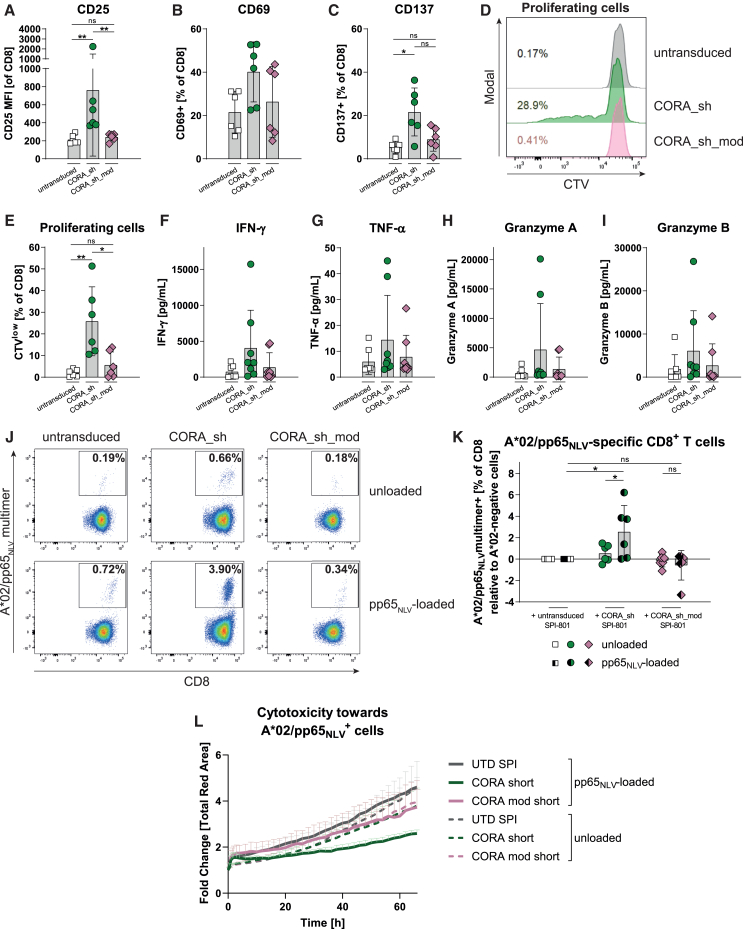
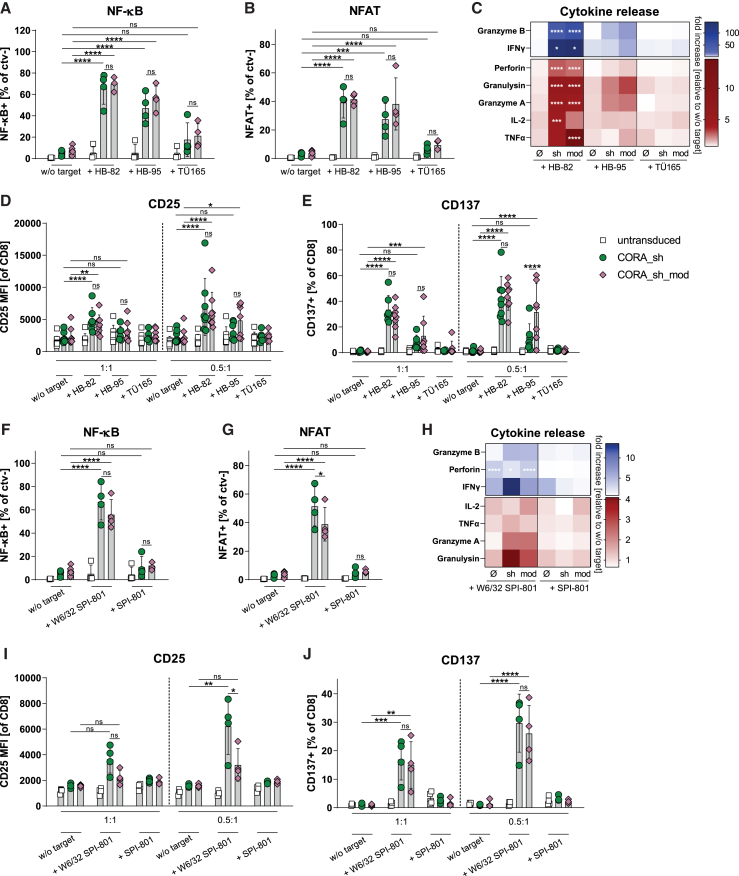
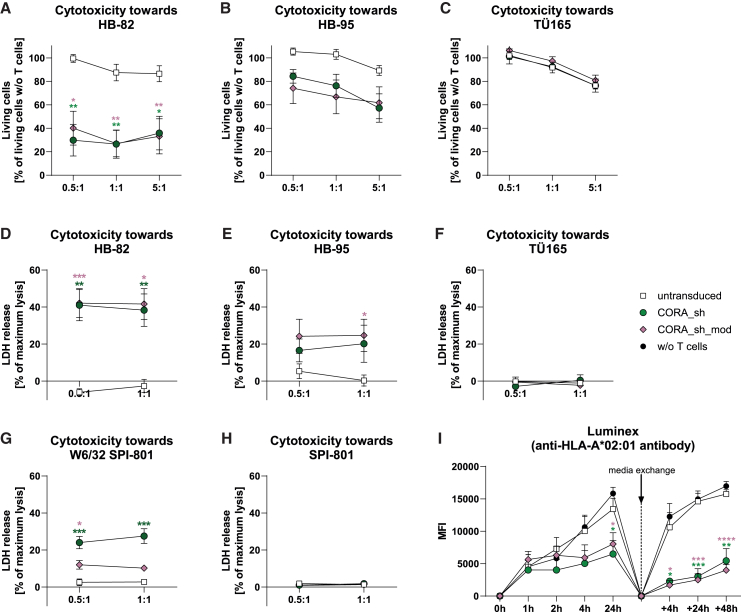
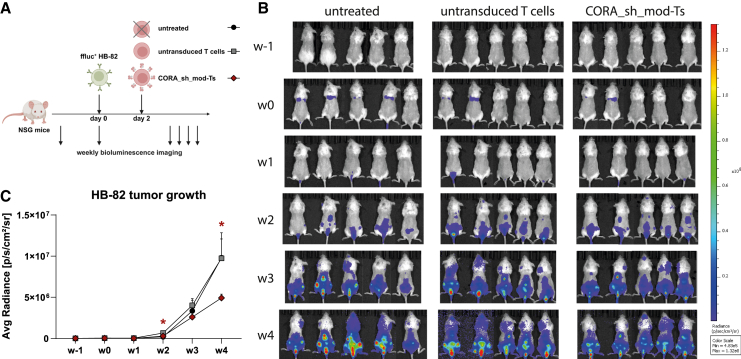
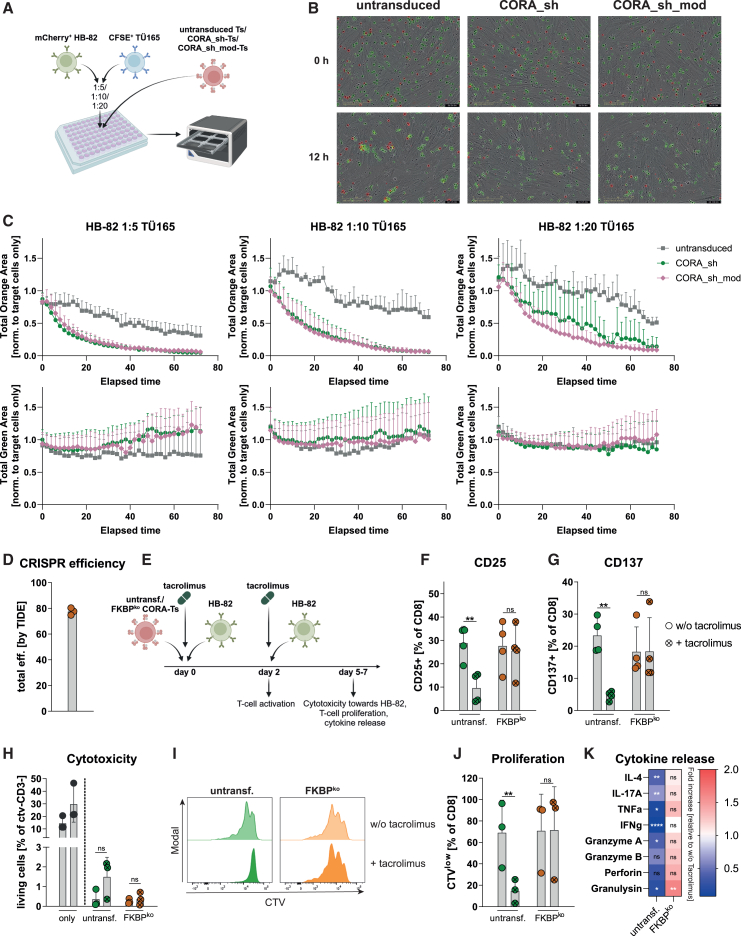
Antibody-mediated rejection (AMR) remains a major complication after solid organ transplantation (SOT). Current treatment options are inefficient and result in drastic impairment of the general immunity. To selectively eliminate responsible alloreactive B cells characterized by anti-donor-HLA B cell receptors (BCRs), we generated T cells overcoming rejection by antibodies (CORA-Ts) engineered with a novel chimeric receptor comprising a truncated donor-HLA molecule as antigen recognition domain. As proof-of-concept, CORA receptors based on HLA-A∗02 were developed. In co-cultures with anti-HLA-A∗02 B cell lines, CORA-Ts were specifically activated, released pro-inflammatory mediators, and exhibited strong cytotoxicity resulting in an effective reduction of anti-HLA-A∗02 antibody release. Significant reduction of growth of an anti-HLA-A∗02 B cell line could be confirmed using an in vivo mouse model. Modification of the CORA receptor effectively abrogated T cell binding, thereby avoiding T cell sensitization. Additionally, using CRISPR-Cas9-mediated knockout of the FKBP12 gene, CORA-Ts were able to resist immunosuppressive treatment with tacrolimus, thereby allowing high efficiency in transplant patients. Our results demonstrate that CORA-Ts are able to specifically eliminate alloreactive, anti-HLA B cells, thus selectively preventing anti-HLA antibody release even under immunosuppressive conditions. This suggests CORA-Ts as potent approach to combat AMR and improve long-term graft survival in SOT patients while preserving their overall B cell immunity.
Keywords: CAR-T cells; HLA; alloreactive B cells; alloreactivity; antibody-mediated rejection; chimeric alloantigen receptor; engineered T cells; mismatch; solid organ transplantation; transplant rejection.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests, except that authors A.C.D., C.F., R.B., and B.E.-V. are inventors of a patent describing the CORA-T approach (EP 3733697, WO 2020/221902: Artificial signaling molecule).
Figures
References
-
- Schlößer H.A., Thelen M., Dieplinger G., von Bergwelt-Baildon A., Garcia-Marquez M., Reuter S., Shimabukuro-Vornhagen A., Wennhold K., Haustein N., Buchner D., et al. Prospective Analyses of Circulating B Cell Subsets in ABO-Compatible and ABO-Incompatible Kidney Transplant Recipients. Am. J. Transpl. 2017;17:542–550. doi: 10.1111/ajt.14013. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
