

Evolution of the RNA alternative decay cis element into a high-affinity target for the immunomodulatory protein Roquin
- PMID: 39801464
- PMCID: PMC11730611
- DOI: 10.1080/15476286.2024.2448391
Evolution of the RNA alternative decay cis element into a high-affinity target for the immunomodulatory protein Roquin
Abstract
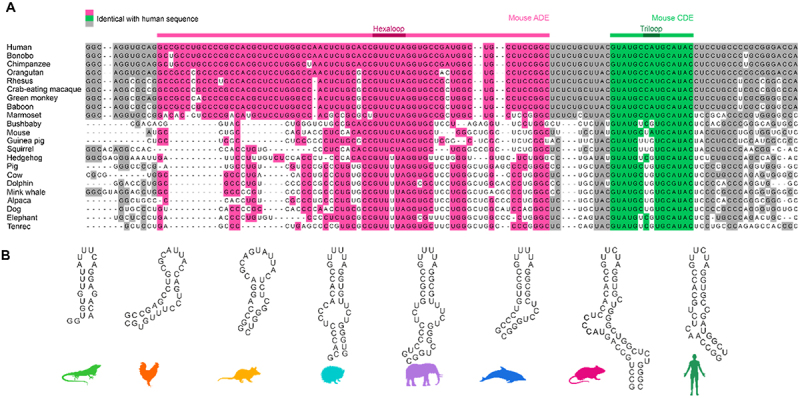
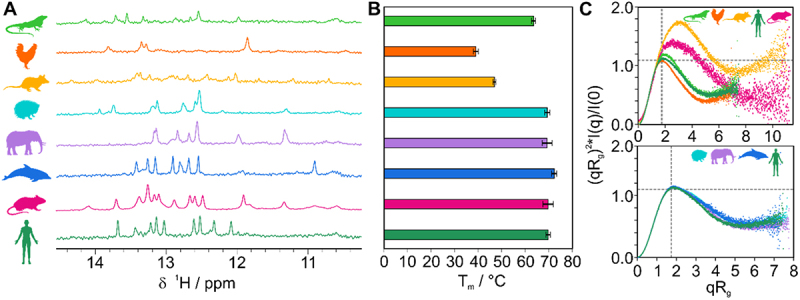
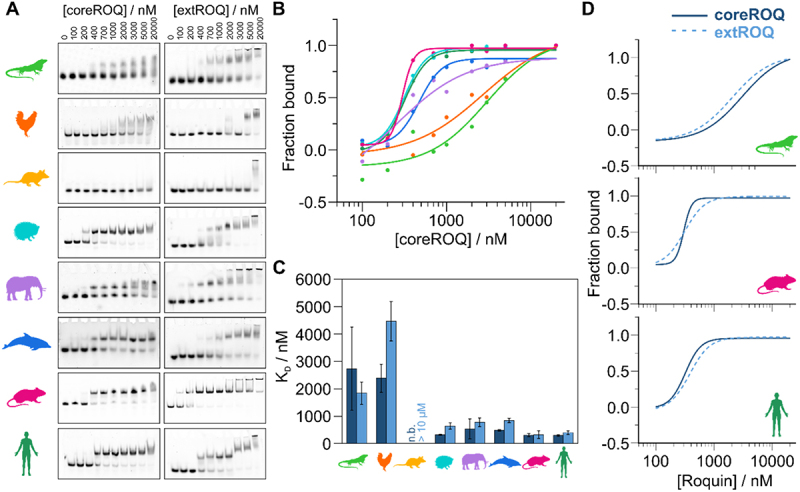
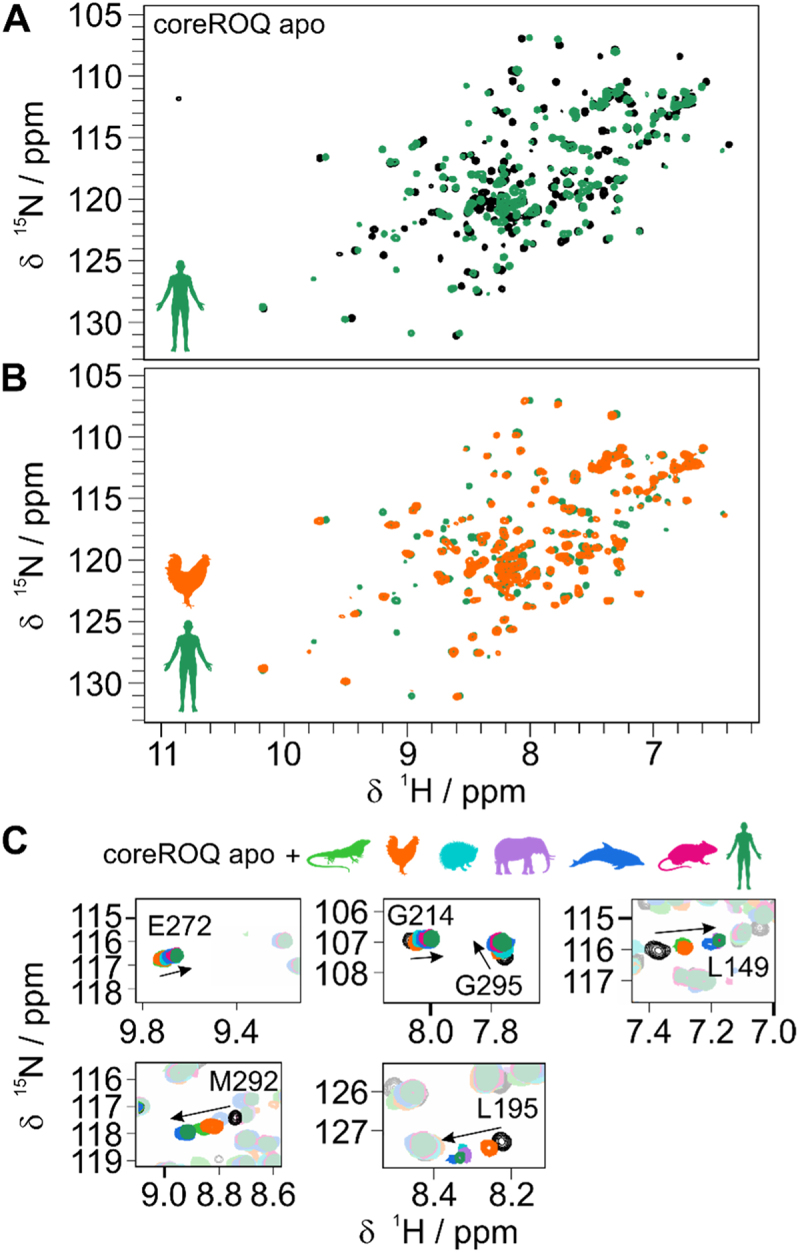
RNA cis elements play pivotal roles in regulatory processes, e.g. in transcriptional and translational regulation. Two stem-looped cis elements, the constitutive and alternative decay elements (CDE and ADE, respectively) are shape-specifically recognized in mRNA 3' untranslated regions (UTRs) by the immune-regulatory protein Roquin. Roquin initiates mRNA decay and contributes to balanced transcript levels required for immune homoeostasis. While the interaction of Roquin with several CDEs is described, our knowledge about ADE complex formation is limited to the mRNA of Ox40, a gene encoding a T-cell costimulatory receptor. The Ox40 3'UTR comprises both a CDE and ADE, each sufficient for Roquin-mediated control. Opposed to highly conserved and abundant CDE structures, ADEs are rarer, but predicted to exhibit a greater structural heterogeneity. This raises the question of how and when two structurally distinct cis elements evolved as equal target motifs for Roquin. Using an interdisciplinary approach, we here monitor the evolution of sequence and structure features of the Ox40 ADE across species. We designed RNA variants to probe en-detail determinants steering Roquin-RNA complex formation. Specifically, those reveal the contribution of a second RNA-binding interface of Roquin for recognition of the ADE basal stem region. In sum, our study sheds light on how the conserved Roquin protein selected ADE-specific structural features to evolve a second high-affinity mRNA target cis element relevant for adaptive immune regulation. As our findings also allow expanding the RNA target spectrum of Roquin, the approach can serve a paradigm for understanding RNA-protein specificity through back-tracing the evolution of the RNA element.
Keywords: EMSA; NMR; RNA; Roquin; SAXS; cis element; geometry; sequence; structure evolution.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
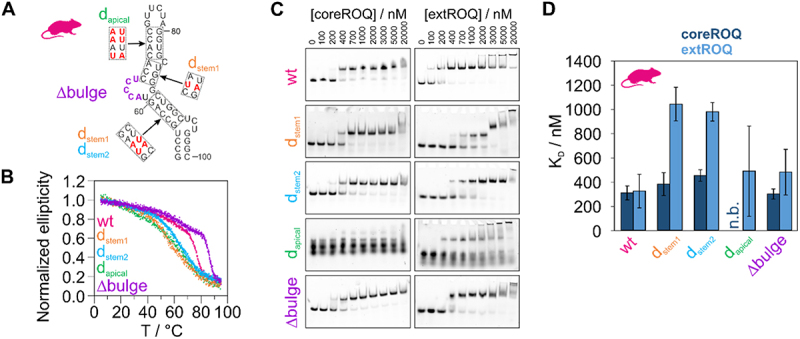
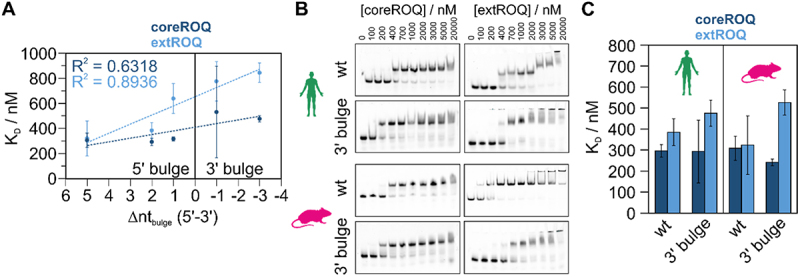
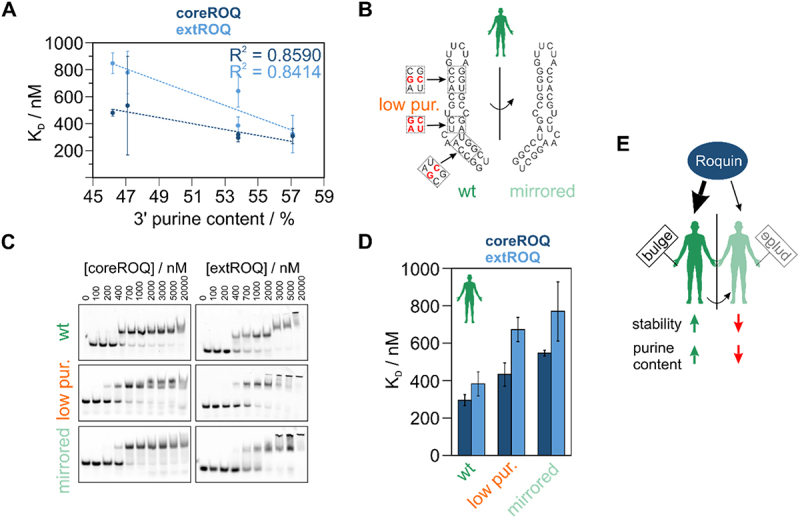
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials