

Ambivalent partnership of the Drosophila posterior class Hox protein Abdominal-B with Extradenticle and Homothorax
- PMID: 39804927
- PMCID: PMC11759358
- DOI: 10.1371/journal.pgen.1011355
Ambivalent partnership of the Drosophila posterior class Hox protein Abdominal-B with Extradenticle and Homothorax
Abstract
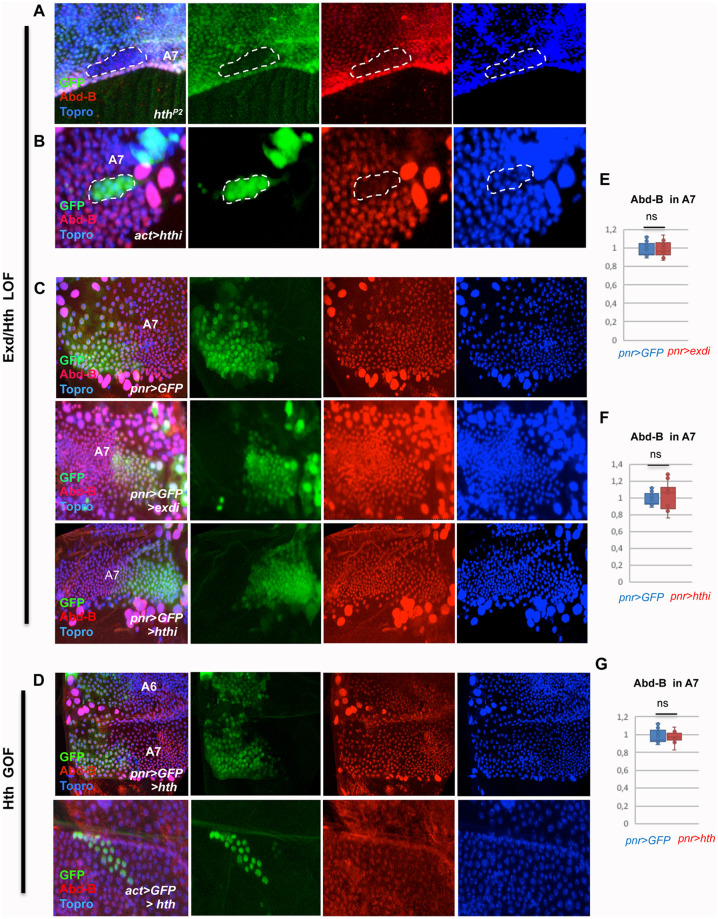
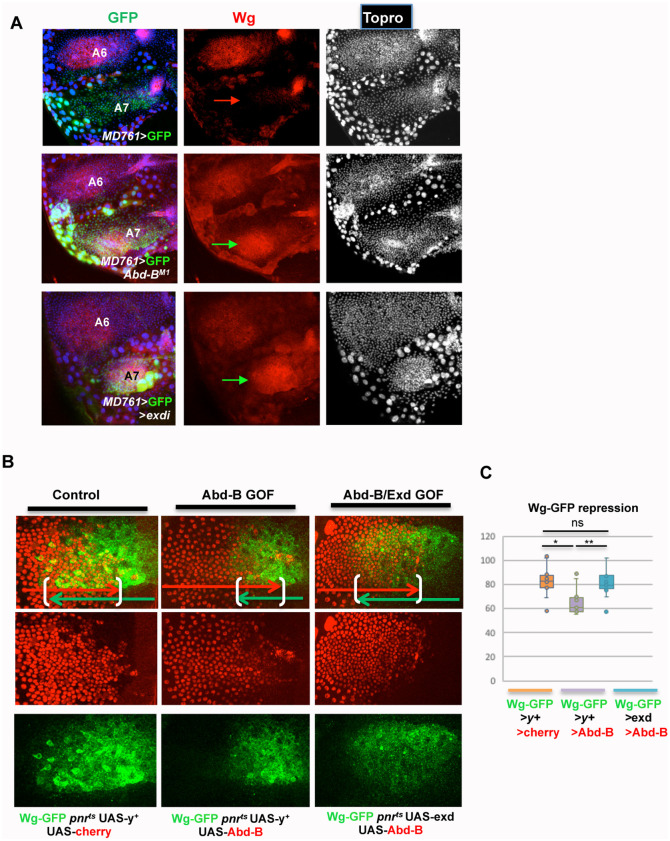
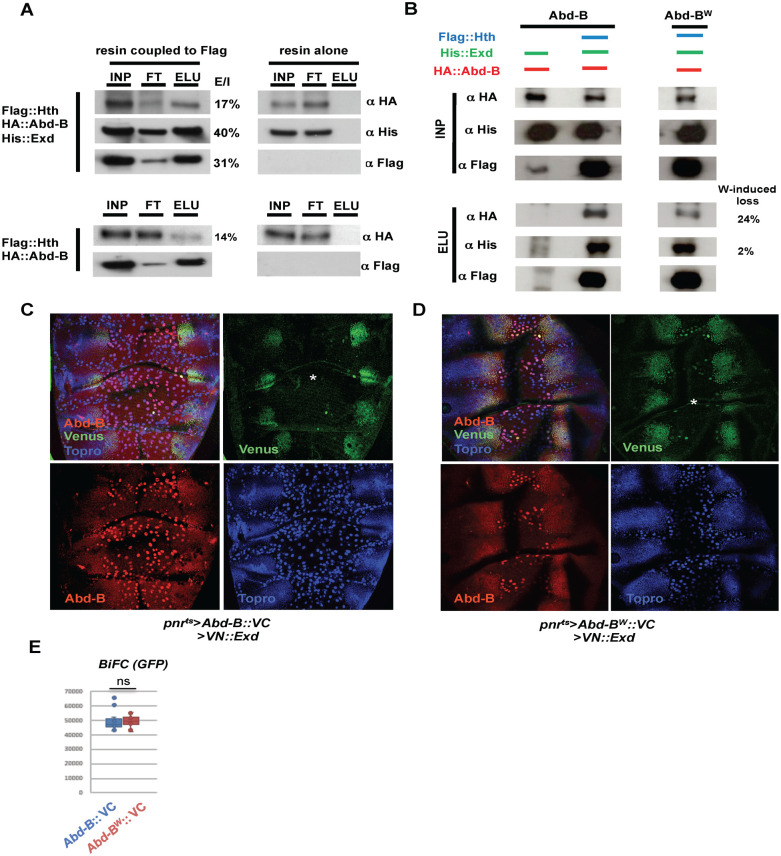
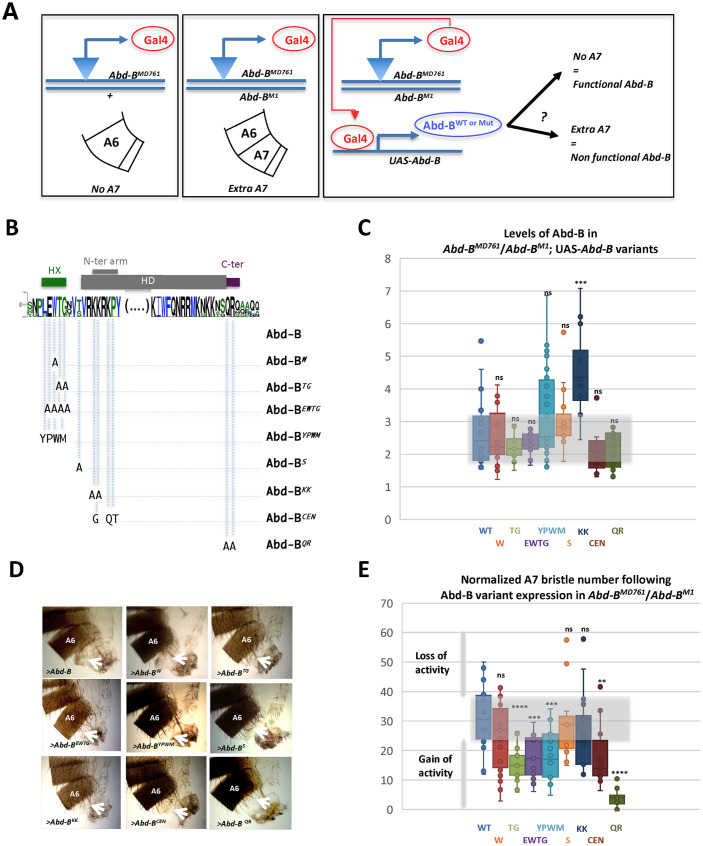
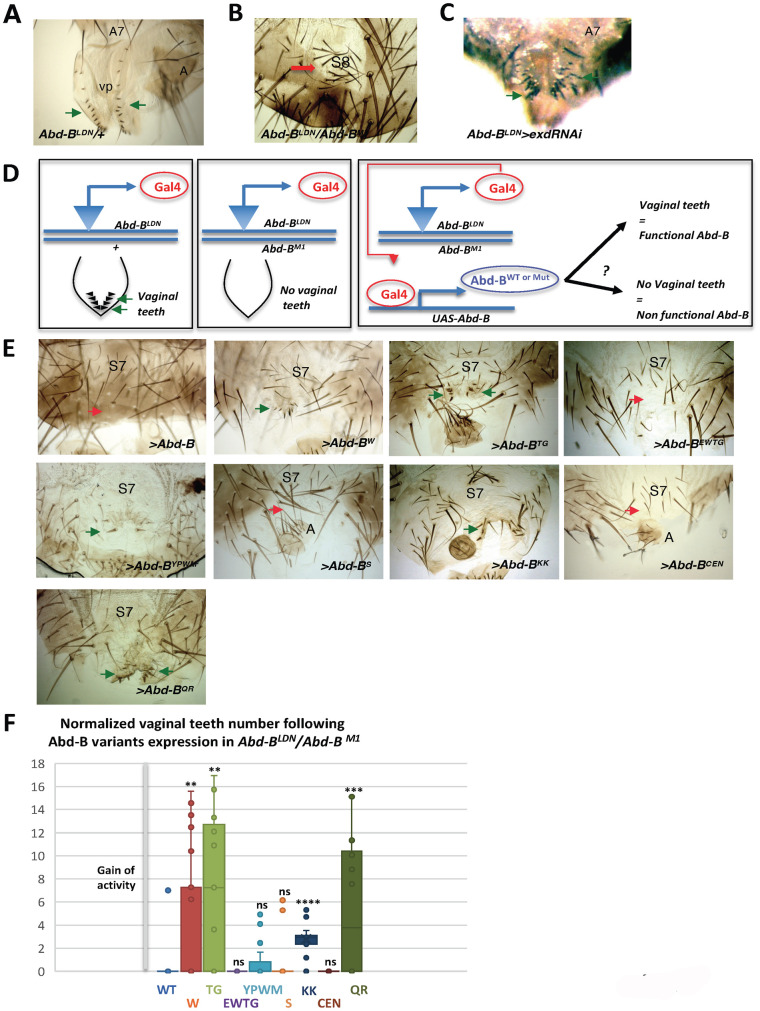
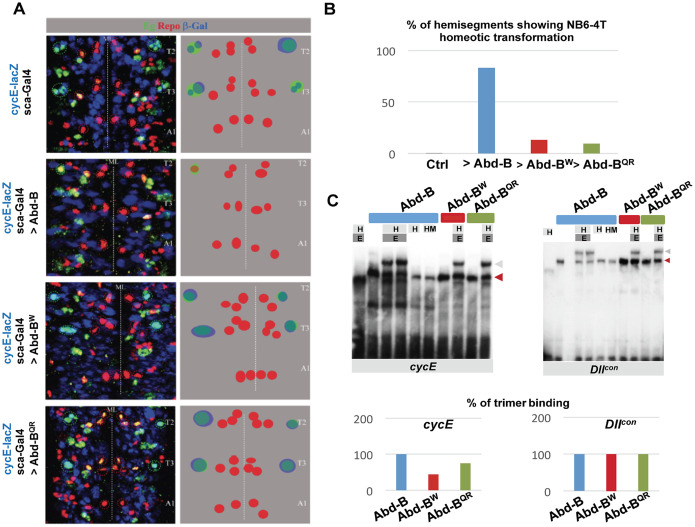
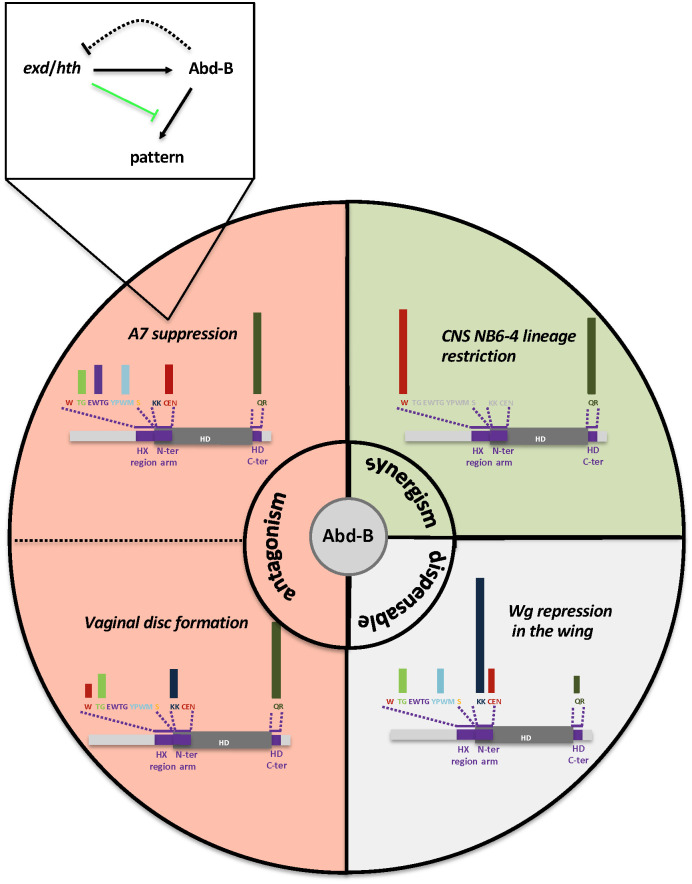
Hox proteins, a sub-group of the homeodomain (HD) transcription factor family, provide positional information for axial patterning in development and evolution. Hox protein functional specificity is reached, at least in part, through interactions with Pbc (Extradenticle (Exd) in Drosophila) and Meis/Prep (Homothorax (Hth) in Drosophila) proteins. Most of our current knowledge of Hox protein specificity stems from the study of anterior and central Hox proteins, identifying the molecular and structural bases for Hox/Pbc/Meis-Prep cooperative action. Posterior Hox class proteins, Abdominal-B (Abd-B) in Drosophila and Hox9-13 in vertebrates, have been comparatively less studied. They strongly diverge from anterior and central class Hox proteins, with a low degree of HD sequence conservation and the absence of a core canonical Pbc interaction motif. Here we explore how Abd-B function interface with that of Exd/Hth using several developmental contexts, studying mutual expression control, functional dependency and intrinsic protein requirements. Results identify cross-regulatory interactions setting relative expression and activity levels required for proper development. They also reveal organ-specific requirement and a binary functional interplay with Exd and Hth, either antagonistic, as previously reported, or synergistic. This highlights context specific use of Exd/Hth, and a similar context specific use of Abd-B intrinsic protein requirements.
Copyright: © 2025 Curt et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Similar articles
-
Antagonism versus cooperativity with TALE cofactors at the base of the functional diversification of Hox protein function.PLoS Genet. 2013;9(2):e1003252. doi: 10.1371/journal.pgen.1003252. Epub 2013 Feb 7. PLoS Genet. 2013. PMID: 23408901 Free PMC article.
-
The cis-regulatory logic underlying abdominal Hox-mediated repression versus activation of regulatory elements in Drosophila.Dev Biol. 2019 Jan 15;445(2):226-236. doi: 10.1016/j.ydbio.2018.11.006. Epub 2018 Nov 20. Dev Biol. 2019. PMID: 30468713 Free PMC article.
-
Hox repression of a target gene: extradenticle-independent, additive action through multiple monomer binding sites.Development. 2002 Jul;129(13):3115-26. doi: 10.1242/dev.129.13.3115. Development. 2002. PMID: 12070087
-
Hox cofactors in vertebrate development.Dev Biol. 2006 Mar 15;291(2):193-206. doi: 10.1016/j.ydbio.2005.10.032. Epub 2006 Mar 3. Dev Biol. 2006. PMID: 16515781 Review.
-
Antennapedia: The complexity of a master developmental transcription factor.Genesis. 2024 Feb;62(1):e23561. doi: 10.1002/dvg.23561. Epub 2023 Oct 13. Genesis. 2024. PMID: 37830148 Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
