

Rice transcription factor bHLH25 confers resistance to multiple diseases by sensing H2O2
- PMID: 39806170
- PMCID: PMC11909244
- DOI: 10.1038/s41422-024-01058-4
Rice transcription factor bHLH25 confers resistance to multiple diseases by sensing H2O2
Abstract
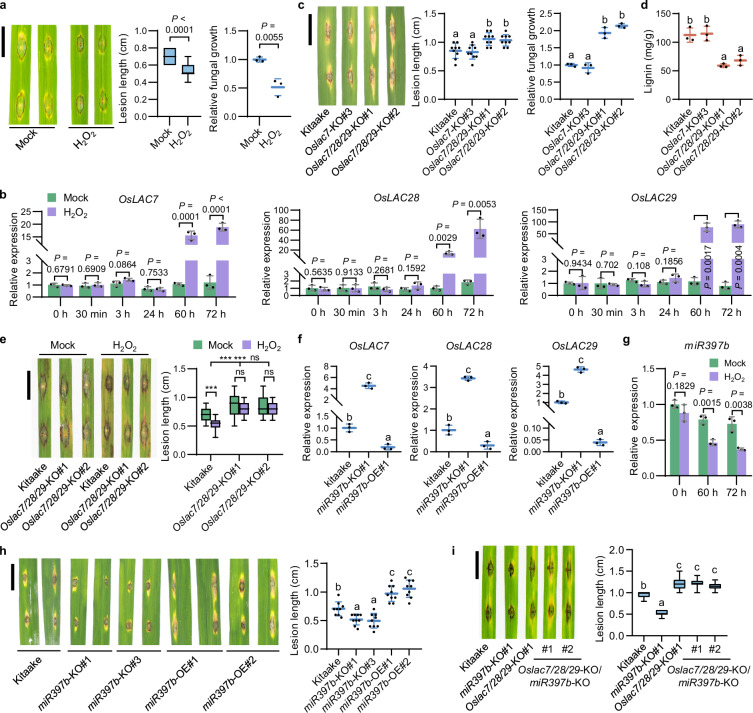
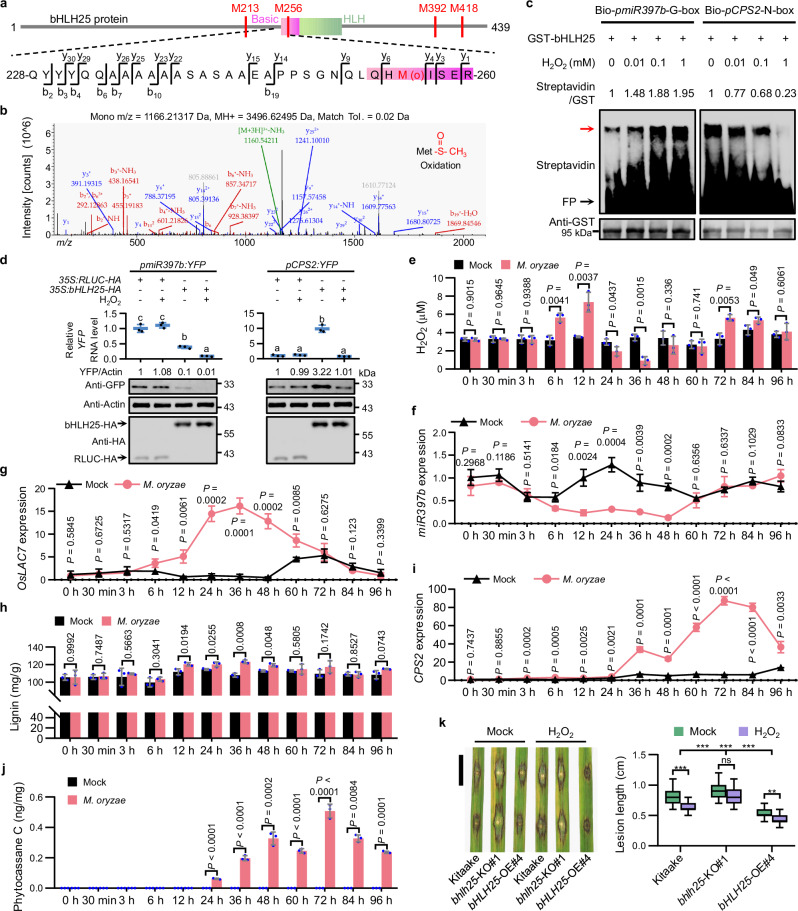
Hydrogen peroxide (H2O2) is a ubiquitous signal regulating many biological processes, including innate immunity, in all eukaryotes. However, it remains largely unknown that how transcription factors directly sense H2O2 in eukaryotes. Here, we report that rice basic/helix-loop-helix transcription factor bHLH25 directly senses H2O2 to confer resistance to multiple diseases caused by fungi or bacteria. Upon pathogen attack, rice plants increase the production of H2O2, which directly oxidizes bHLH25 at methionine 256 in the nucleus. Oxidized bHLH25 represses miR397b expression to activate lignin biosynthesis for plant cell wall reinforcement, preventing pathogens from penetrating plant cells. Lignin biosynthesis consumes H2O2 causing accumulation of non-oxidized bHLH25. Non-oxidized bHLH25 switches to promote the expression of Copalyl Diphosphate Synthase 2 (CPS2), which increases phytoalexin biosynthesis to inhibit expansion of pathogens that escape into plants. This oxidization/non-oxidation status change of bHLH25 allows plants to maintain H2O2, lignin and phytoalexin at optimized levels to effectively fight against pathogens and prevents these three molecules from over-accumulation that harms plants. Thus, our discovery reveals a novel mechanism by which a single protein promotes two independent defense pathways against pathogens. Importantly, the bHLH25 orthologues from available plant genomes all contain a conserved M256-like methionine suggesting the broad existence of this mechanism in the plant kingdom. Moreover, this Met-oxidation mechanism may also be employed by other eukaryotic transcription factors to sense H2O2 to change functions.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures




References
-
- Sundaresan, M., Yu, Z. X., Ferrans, V. J., Irani, K. & Finkel, T. Requirement for generation of H2O2 for platelet-derived growth factor signal transduction. Science270, 296–299 (1995). - PubMed
-
- Sies, H. & Jones, D. P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol.21, 363–383 (2020). - PubMed
-
- Mittler, R., Zandalinas, S. I., Fichman, Y. & Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol.23, 663–679 (2022). - PubMed
MeSH terms
Substances
Grants and funding
- 32425005/National Natural Science Foundation of China (National Science Foundation of China)
- 32072043/National Natural Science Foundation of China (National Science Foundation of China)
- 2023NSFSC0005/Department of Science and Technology of Sichuan Province (Sichuan Provincial Department of Science and Technology)
LinkOut - more resources
Full Text Sources
Miscellaneous
