

Stepwise order in protein complex assembly: approaches and emerging themes
- PMID: 39809320
- PMCID: PMC11732423
- DOI: 10.1098/rsob.240283
Stepwise order in protein complex assembly: approaches and emerging themes
Abstract
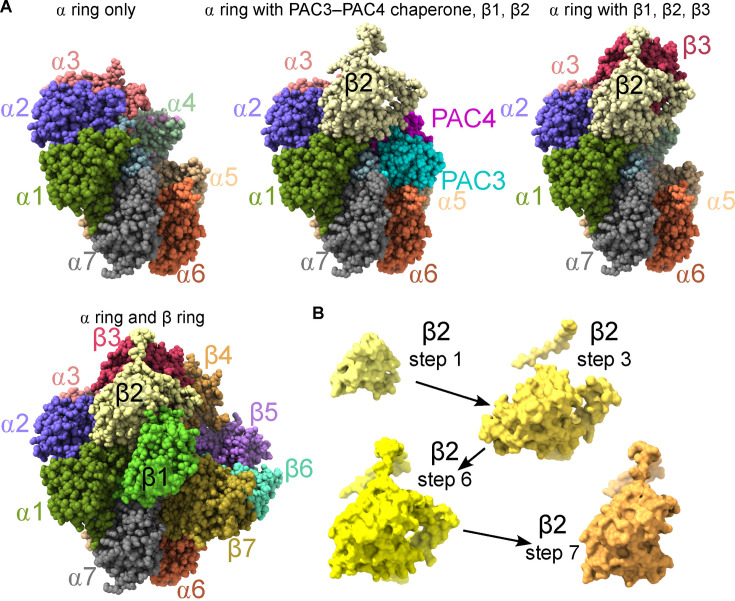
Protein-based nanomachines drive every cellular process. An explosion of high-resolution structures of multiprotein complexes has improved our understanding of what these machines look like and how they work, but we still know relatively little about how they assemble in living cells. For example, it has only recently been appreciated that many complexes assemble co-translationally, with at least one subunit still undergoing active translation while already interacting with other subunits. One aspect that is particularly understudied is assembly order, the idea that there is a stepwise order to the subunit-subunit associations that underlies the efficient assembly of the quaternary structure. Here, we integrate a review of the methodological approaches commonly used to query assembly order within a discussion of studies of the 20S proteasome core particle, septin protein complexes, and the histone octamer. We highlight shared and distinct properties of these complexes that illustrate general themes applicable to most other multisubunit assemblies.
Keywords: assembly; oligomerization; order; protein complexes; subunits.
Conflict of interest statement
We declare we have no competing interests.
Figures




References
-
- Levinthal C. 1969. Mossbauer spectroscopy in biological systems (eds Debrunner P, Tsibris JCM, Munck E). In Proceedings of a meeting held at Allerton House. Urbana, IL: University of Illinois Press.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
