

The small GTPase MRAS is a broken switch
- PMID: 39809765
- PMCID: PMC11733253
- DOI: 10.1038/s41467-025-55967-y
The small GTPase MRAS is a broken switch
Abstract
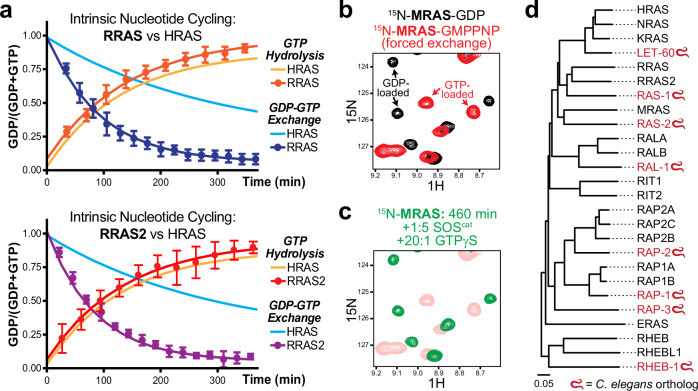
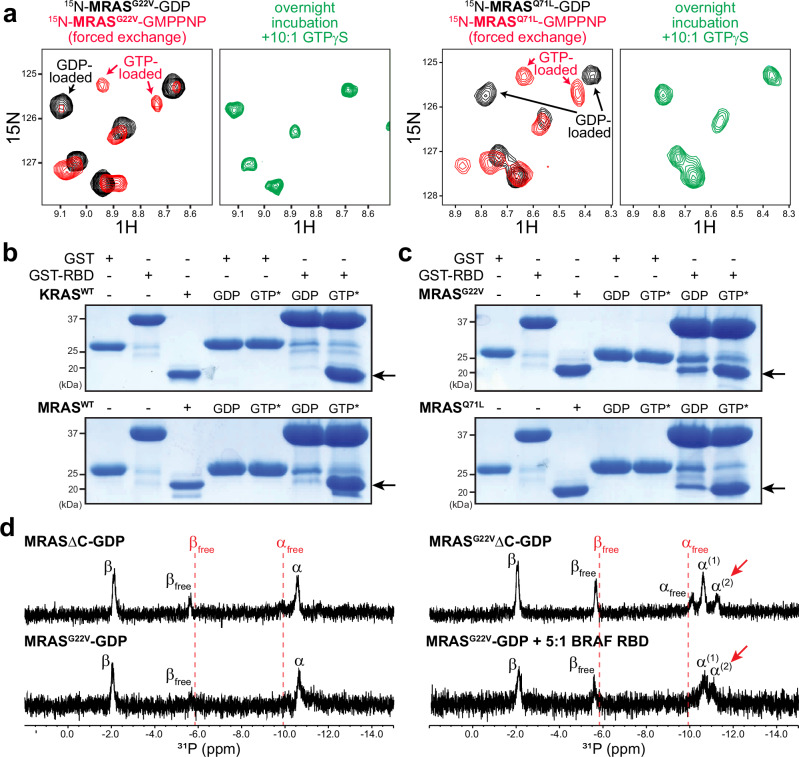
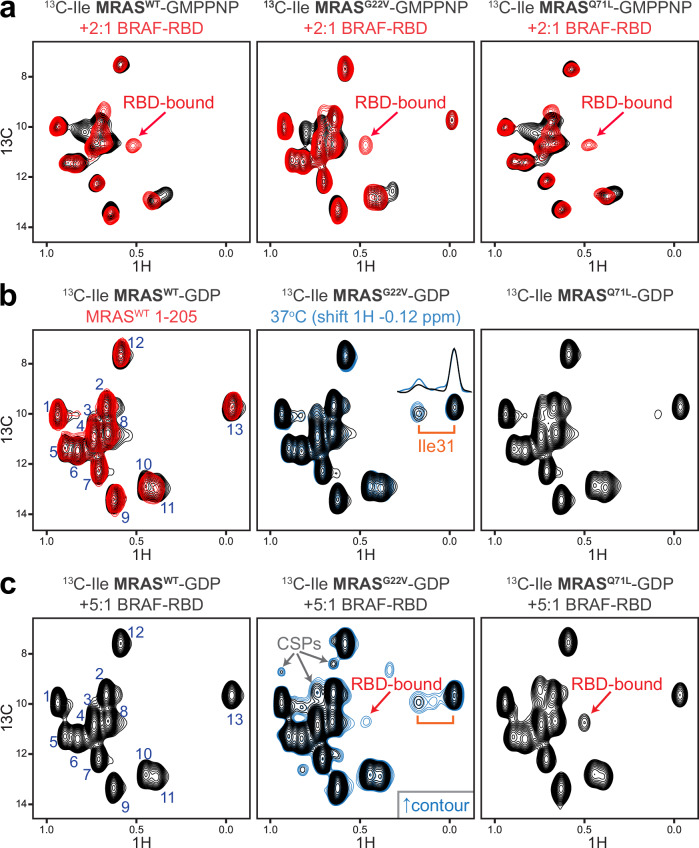
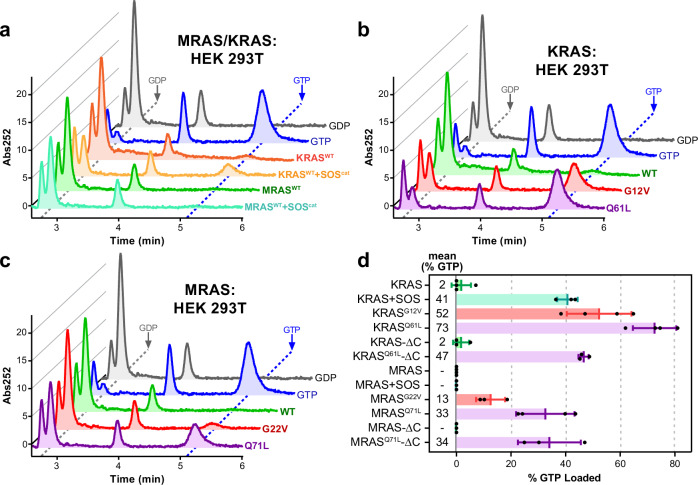
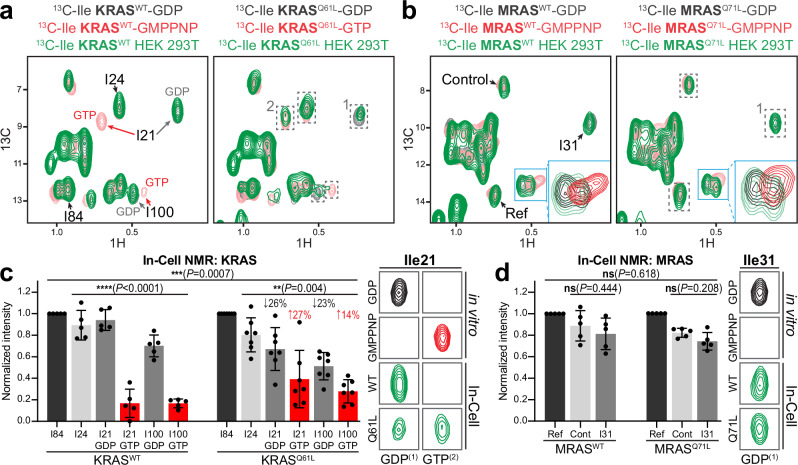
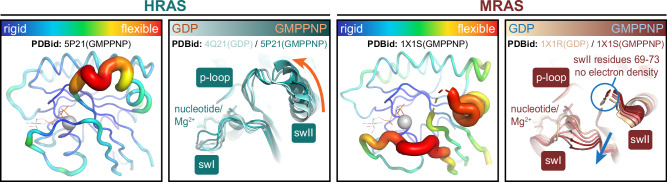
Intense research on founding members of the RAS superfamily has defined our understanding of these critical signalling proteins, leading to the premise that small GTPases function as molecular switches dependent on differential nucleotide loading. The closest homologs of H/K/NRAS are the three-member RRAS family, and interest in the MRAS GTPase as a regulator of MAPK activity has recently intensified. We show here that MRAS does not function as a classical switch and is unable to exchange GDP-to-GTP in solution or when tethered to a lipid bilayer. The exchange defect is unaffected by inclusion of the GEF SOS1 and is conserved in a distal ortholog from nematodes. Synthetic activating mutations widely used to study the function of MRAS in a presumed GTP-loaded state do not increase exchange, but instead drive effector binding due to sampling of an activated conformation in the GDP-loaded state. This includes nucleation of the SHOC2-PP1Cα holophosphatase complex. Acquisition of NMR spectra from isotopically labeled MRAS in live cells validated the GTPase remains fully GDP-loaded, even a supposed activated mutant. These data show that RAS GTPases, including those most similar to KRAS, have disparate biochemical activities and challenge current dogma on MRAS, suggesting previous data may need reinterpretation.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures


References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
