

Enabling next-generation engineered TCR-T therapies based on high-throughput TCR discovery from diagnostic tumor biopsies
- PMID: 39809767
- PMCID: PMC11733228
- DOI: 10.1038/s41467-024-55420-6
Enabling next-generation engineered TCR-T therapies based on high-throughput TCR discovery from diagnostic tumor biopsies
Abstract
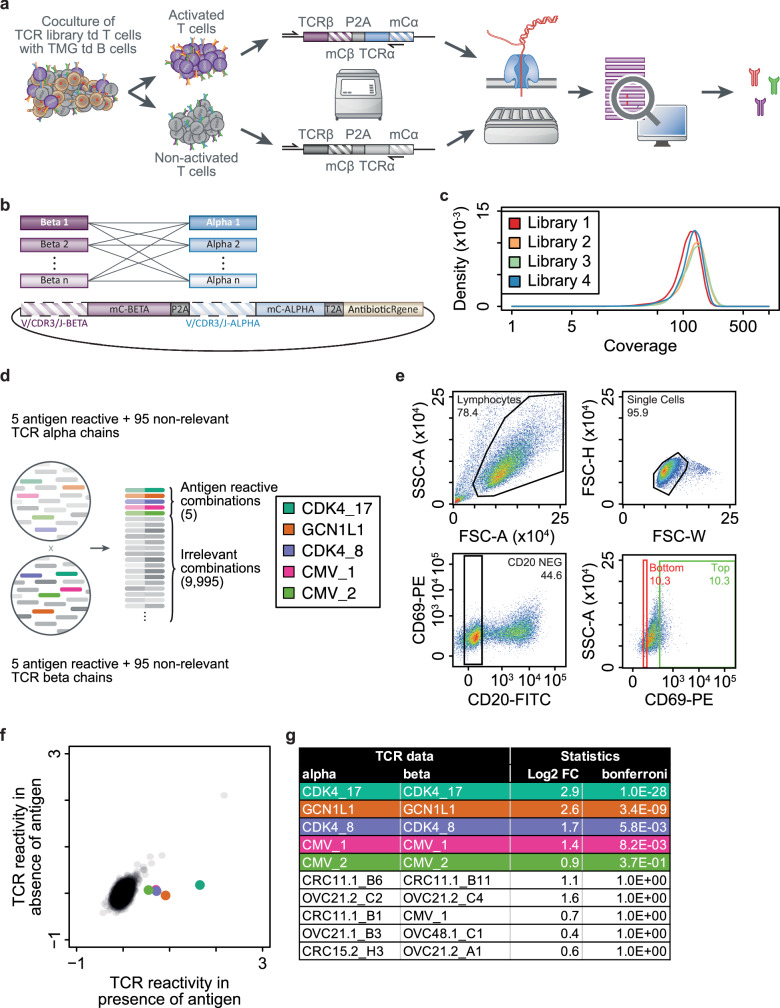
Adoptive cell therapy with tumor-infiltrating lymphocytes (TIL) can mediate tumor regression, including complete and durable responses, in a range of solid cancers, most notably in melanoma. However, its wider application and efficacy has been restricted by the limited accessibility, proliferative capacity and effector function of tumor-specific TIL. Here, we develop a platform for the efficient identification of tumor-specific TCR genes from diagnostic tumor biopsies, including core-needle biopsies frozen in a non-viable format, to enable engineered T cell therapy. Using a genetic screening approach that detects antigen-reactive TCRs with high sensitivity and specificity based on T cell activation, we show that high complexity TCR libraries can be efficiently screened against multiplexed antigen libraries to identify both HLA class I and II restricted TCRs. Through the identification of neoantigen-specific TCRs directly from melanoma as well as low tumor mutational burden microsatellite-stable colorectal carcinoma samples, we demonstrate the pan-cancer potential of this platform.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: T.K., D.S.S., J.G., R.G., R.M.S., L.B., J.W., H.K., D.H., M.V., Y.B.C., M.Saornil., O.K., B.S., H.D., A.G., A.C.M., B.W., M.L., C.G.E., M.Sabatino., J.v.d.B., J.W.J.v.H., G.M.B., and C.L. are salaried employees and stock option holders of Neogene Therapeutics. K.Y.T is a compensated consultant for Verrica Pharmaceuticals and NFlection Therapeutics (also with ownership interest), and receives research support from Incyte Corporation. J.J.M. is Associate Center Director at Moffitt Cancer Center, has ownership interest in Aleta Biotherapeutics, CG Oncology, Turnstone Biologics, Ankyra Therapeutics, and AffyImmune Therapeutics, and is a paid consultant/paid advisory board member for ONCoPEP, CG Oncology, Mersana Therapeutics, Turnstone Biologics, Vault Pharma, Ankyra Therapeutics, AffyImmune Therapeutics, UbiVac, Vycellix, and Aleta Biotherapeutics. V.K.S. is a compensated consultant for Alkermes, Bristol Myers Squibb, Genesis Drug Discovery & Development, Iovance, Merck, Mural Oncology, and Novartis, and receives research funding from SkylineDX and Turnstone, in addition to Neogene Therapeutics. I.J. is involved in projects supported by research agreements between the NKI and Neogene Therapeutics, Asher Biotherapeutics and Sastra Cell Therapy. J.H, is a member of the Neogene Therapeutics Scientific Advisory Board and is a stock option holder of Neogene Therapeutics. He is also advisor for Achilles Therapeutics, BioNTech, Instil Bio, PokeAcell, Scenic Biotech, T-Knife and Third Rock Ventures. T.N.S. is co-founder of Neogene Therapeutics; is advisor for Allogene Therapeutics, Asher Bio, Celsius, Merus, Neogene Therapeutics, and Scenic Biotech; is a stockholder in Allogene Therapeutics, Asher Bio, Cell Control, Celsius, Merus, Neogene Therapeutics and Scenic Biotech; and is venture partner at Third Rock Ventures. The remaining authors declare no competing interests. The Netherlands Cancer Institute has entered into a clinical trial collaboration with Neogene Therapeutics. The TCR library screening technology is described in patents WO2021011482A1 and US2021040558A1 (pending) with inventors C.L., T.N.S., D.S.S., T.K., J.W.J.v.H. G.M.B., R.G. and J.G. and the B cell immortalization technology is described in patent US20220228164A1 (pending) with inventors T.N.S, C.L., T.K., G.M.B., J.G., J.W.J.v.H., R.G., D.S.S., J.W. and L.B.; all patents are assigned to Neogene Therapeutics B.V.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
