

Splicing to orchestrate cell fate
- PMID: 39811494
- PMCID: PMC11729663
- DOI: 10.1016/j.omtn.2024.102416
Splicing to orchestrate cell fate
Abstract
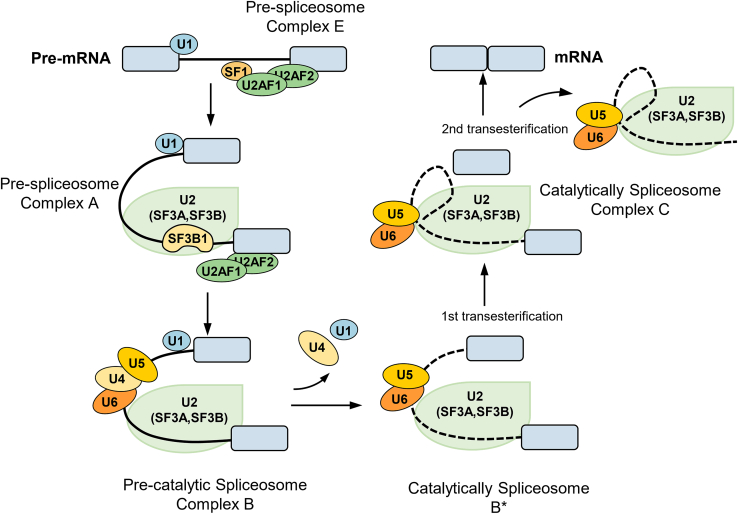
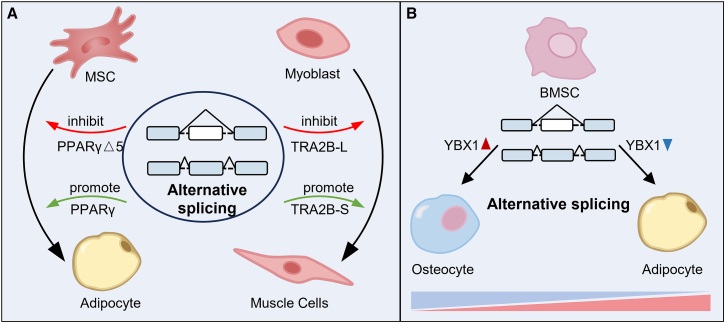
Alternative splicing (AS) plays a critical role in gene expression by generating protein diversity from single genes. This review provides an overview of the role of AS in regulating cell fate, focusing on its involvement in processes such as cell proliferation, differentiation, apoptosis, and tumorigenesis. We explore how AS influences the cell cycle, particularly its impact on key stages like G1, S, and G2/M. The review also examines AS in cell differentiation, highlighting its effects on mesenchymal stem cells and neurogenesis, and how it regulates differentiation into adipocytes, osteoblasts, and chondrocytes. Additionally, we discuss the role of AS in programmed cell death, including apoptosis and pyroptosis, and its contribution to cancer progression. Importantly, targeting aberrant splicing mechanisms presents promising therapeutic opportunities for restoring normal cellular function. By synthesizing recent findings, this review provides insights into how AS governs cellular fate and offers directions for future research into splicing regulatory networks.
Keywords: MT: Bioinformatics; alternative splicing; apoptosis; cell cycle; cell differentiation; neurogenesis; tumorigenesis.
© 2024 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Chow L.T., Gelinas R.E., Broker T.R., Roberts R.J. An amazing sequence arrangement at the 5' ends of adenovirus 2 messenger RNA. Cell. 1977;12:1–8. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Research Materials
