

De novo designed proteins neutralize lethal snake venom toxins
- PMID: 39814879
- PMCID: PMC11882462
- DOI: 10.1038/s41586-024-08393-x
De novo designed proteins neutralize lethal snake venom toxins
Abstract
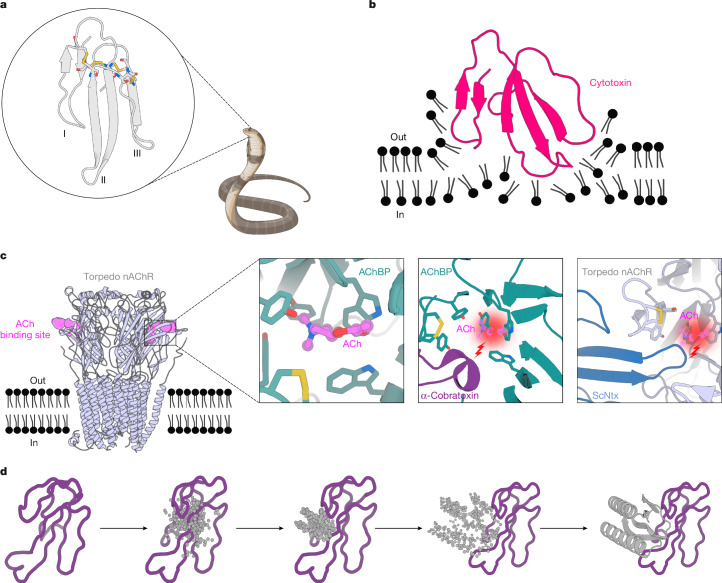
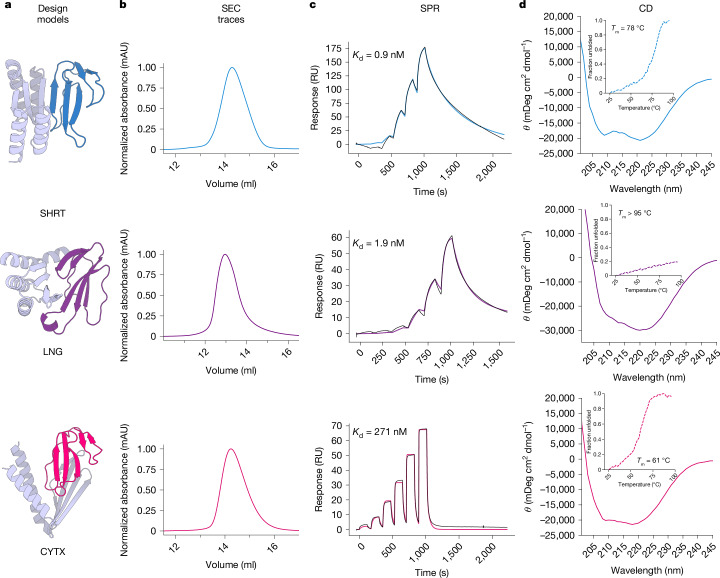
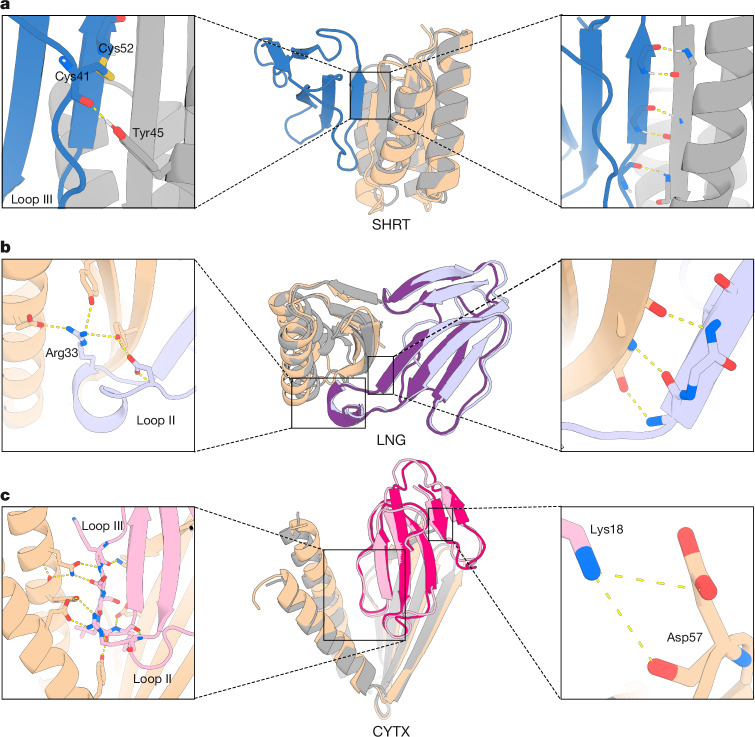
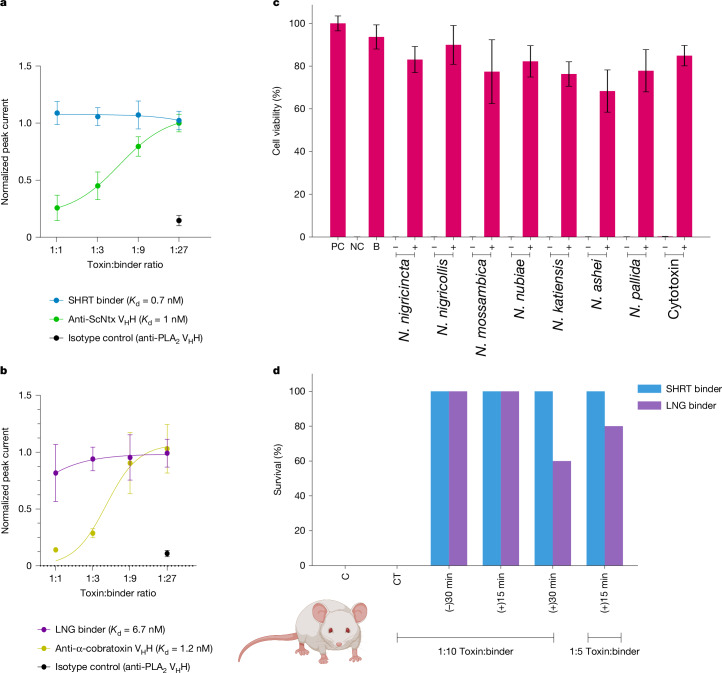
Snakebite envenoming remains a devastating and neglected tropical disease, claiming over 100,000 lives annually and causing severe complications and long-lasting disabilities for many more1,2. Three-finger toxins (3FTx) are highly toxic components of elapid snake venoms that can cause diverse pathologies, including severe tissue damage3 and inhibition of nicotinic acetylcholine receptors, resulting in life-threatening neurotoxicity4. At present, the only available treatments for snakebites consist of polyclonal antibodies derived from the plasma of immunized animals, which have high cost and limited efficacy against 3FTxs5-7. Here we used deep learning methods to de novo design proteins to bind short-chain and long-chain α-neurotoxins and cytotoxins from the 3FTx family. With limited experimental screening, we obtained protein designs with remarkable thermal stability, high binding affinity and near-atomic-level agreement with the computational models. The designed proteins effectively neutralized all three 3FTx subfamilies in vitro and protected mice from a lethal neurotoxin challenge. Such potent, stable and readily manufacturable toxin-neutralizing proteins could provide the basis for safer, cost-effective and widely accessible next-generation antivenom therapeutics. Beyond snakebite, our results highlight how computational design could help democratize therapeutic discovery, particularly in resource-limited settings, by substantially reducing costs and resource requirements for the development of therapies for neglected tropical diseases.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: D.B., S.V.T., T.P.J., M.B.V., S.P.M., S.K.M., N.R.C., E.M., I.S., A.K.B. and A.K. are inventors on a provisional patent application submitted by the University of Washington for the design and composition of the proteins created in this study.
Figures
Update of
-
De novo designed proteins neutralize lethal snake venom toxins.Res Sq [Preprint]. 2024 May 17:rs.3.rs-4402792. doi: 10.21203/rs.3.rs-4402792/v1. Res Sq. 2024. Update in: Nature. 2025 Mar;639(8053):225-231. doi: 10.1038/s41586-024-08393-x. PMID: 38798548 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
