

Inducible engineering precursor metabolic flux for synthesizing hyaluronic acid of customized molecular weight in Streptococcus zooepidemicus
- PMID: 39825423
- PMCID: PMC11748608
- DOI: 10.1186/s12934-024-02624-6
Inducible engineering precursor metabolic flux for synthesizing hyaluronic acid of customized molecular weight in Streptococcus zooepidemicus
Abstract
Background: Hyaluronic acid (HA) is extensively employed in various fields such as medicine, cosmetics, food, etc. The molecular weight (MW) of HA is crucial for its biological functions. Streptococcus zooepidemicus, a prominent HA industrial producer, naturally synthetizes HA with high MW. Currently, few effective approaches exist for the direct and precise regulation of HA MW through a one-step fermentation process, and S. zooepidemicus lacks metabolic regulatory elements with varying intensities. The ratio of HA's precursors, UDP-N-acetylglucosamine (UDP-GlcNAc) and UDP-glucuronic acid (UDP-GlcA), is critical for the extension and release of HA. An imbalance in the precursor proportions for HA synthesis leads to a significant decrease in HA MW, indicating that controlling the precursor ratio may serve as a potential method for regulating HA MW.
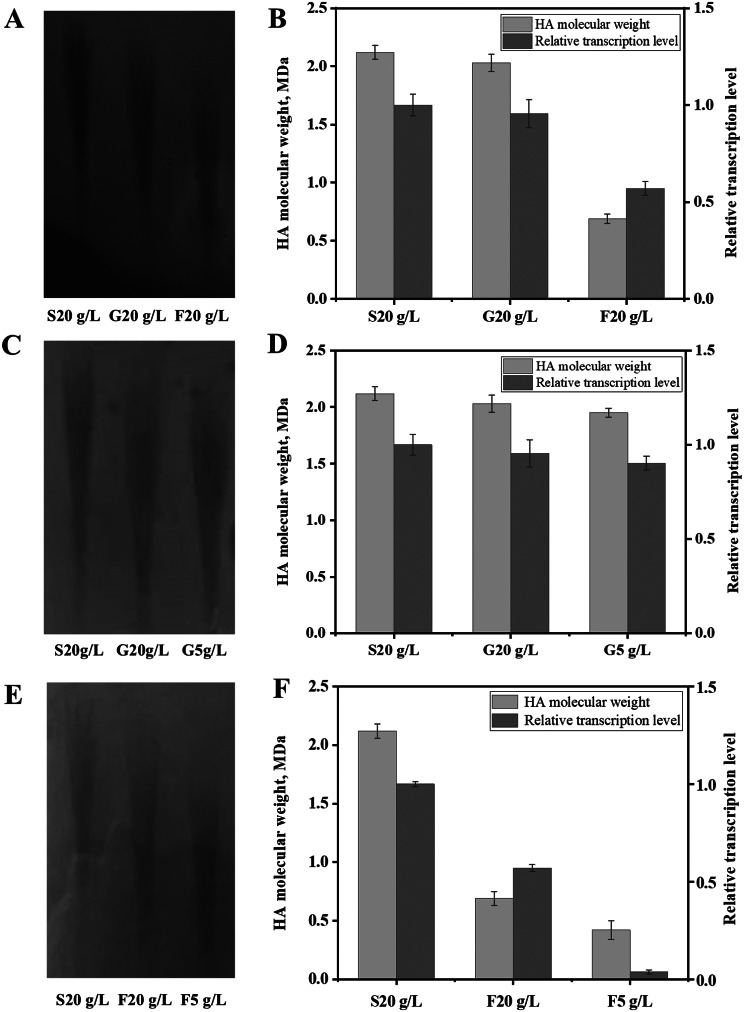
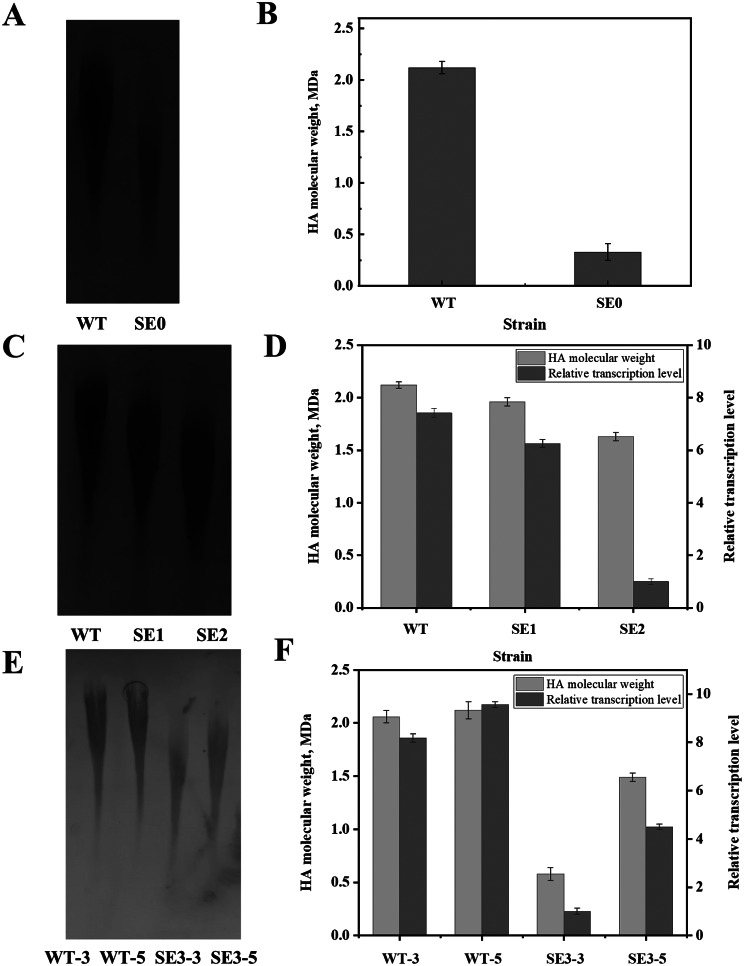
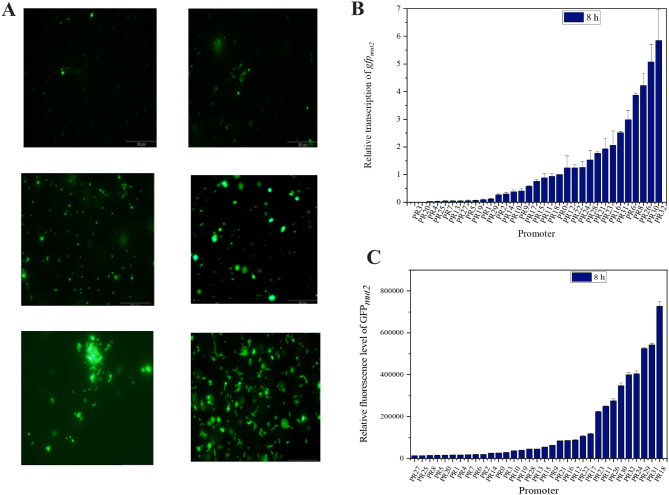
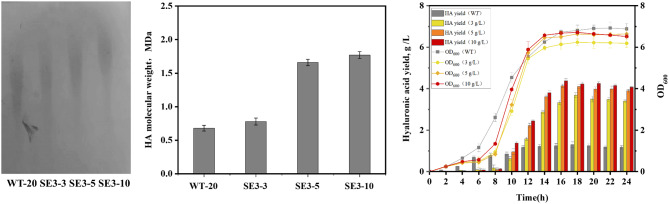
Results: In this study, the type and concentration of carbon sources were manipulated to disrupt the balance of precursor supply. Based on the results, it was speculated that the transcription level of hasE, which may connect the two HA synthesis precursors, is positively correlated with HA MW. Consequently, an endogenous expression component library for S. zooepidemicus was constructed, comprising 32 constitutive and 4 inducible expression elements. The expression of hasE was subsequently regulated in strain SE0 (S12 ΔhasE) using two constitutive promoters of differing strengths. The recombinant strain SE1, in which hasE was controlled by the stronger promoter PR31, produced HA with a MW of 1.96 MDa. In contrast, SE2, utilizing the weaker promoter PR22, synthesized shorter HA with a MW of 1.63 MDa, thereby verifying the hypothesis. Finally, to precisely regulate HA MW according to specific demands, an efficient sucrose-induced expression system was screened and employed to control the transcription level of hasE, obtaining recombinant strain SE3. When induced with sucrose concentrations of 3, 5-10 g/L, the HA MW of SE3 reached 0.78 to 1.77 MDa, respectively.
Conclusions: Studies on regulating the balance of the HA precursor substances indicate that an oversupply of either UDP-GlcNAc or UDP-GlcUA can reduce HA MW. The hasE gene serves as a crucial regulator for maintaining this balance. Precise regulation of hasE transcription was achieved through an efficient inducible expression system, enabling the customized production of HA with specific MW. The HA MW of strain SE3 can be accurately manipulated by adjusting sucrose concentration, establishing a novel strategy for customized HA fermentation.
Keywords: Streptococcus zooepidemicus; hasE gene; Hyaluronic acid; Molecular weight regulation; Precursor supply balance; Sucrose-induced expression system.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures


Similar articles
-
The role of hyaluronic acid precursor concentrations in molecular weight control in Streptococcus zooepidemicus.Mol Biotechnol. 2014 Feb;56(2):147-56. doi: 10.1007/s12033-013-9690-4. Mol Biotechnol. 2014. PMID: 23903961
-
Hyaluronan molecular weight is controlled by UDP-N-acetylglucosamine concentration in Streptococcus zooepidemicus.J Biol Chem. 2009 Jul 3;284(27):18007-14. doi: 10.1074/jbc.M109.011999. Epub 2009 May 18. J Biol Chem. 2009. PMID: 19451654 Free PMC article.
-
Production of controlled molecular weight hyaluronic acid by glucostat strategy using recombinant Lactococcus lactis cultures.Appl Microbiol Biotechnol. 2019 Jun;103(11):4363-4375. doi: 10.1007/s00253-019-09769-0. Epub 2019 Apr 9. Appl Microbiol Biotechnol. 2019. PMID: 30968163
-
Genetic basis for hyper production of hyaluronic acid in natural and engineered microorganisms.Microb Cell Fact. 2016 Jul 1;15(1):119. doi: 10.1186/s12934-016-0517-4. Microb Cell Fact. 2016. PMID: 27370777 Free PMC article. Review.
-
Microbial production of hyaluronic acid: current state, challenges, and perspectives.Microb Cell Fact. 2011 Nov 16;10:99. doi: 10.1186/1475-2859-10-99. Microb Cell Fact. 2011. PMID: 22088095 Free PMC article. Review.
Cited by
-
Hyaluronic Acid and Its Synthases-Current Knowledge.Int J Mol Sci. 2025 Jul 22;26(15):7028. doi: 10.3390/ijms26157028. Int J Mol Sci. 2025. PMID: 40806161 Free PMC article. Review.
References
-
- Stern R, Asari AA, Sugahara K. Hyaluronan fragments: an information-rich system. Cell Biol. 2006;85(8):699–715. 10.1016/j.ejcb.2006.05.009. - PubMed
-
- Graça MFP, Miguel SP, Cabral CSD, Correia IJ. Hyaluronic acid-based wound dressings: a review. Carbohydr Polym. 2020;241:116364–417. 10.1016/j.carbpol.2020.116364. - PubMed
-
- Diane R, Leonardo M, Silva D, Fischer B. Hyaluronic acid – from production to application: a review. BRIAC. 2023;13(3):211–33. 10.33263/BRIAC133.211.
-
- Zheng XL, Wang BT, Tang X, Mao BY, Zhang QX, Zhang TM, Zhao JX, Cui SM, Chen W. Absorption, metabolism, and functions of HA and its therapeutic prospects in combination with microorganisms: A review. Carbohydr Polym. 2023; 299:120153-71. 10.1016/ j. carbpol. 2022. 120153. - PubMed
-
- Johnson ME, Murphy PJ, Boulton M. Effectiveness of sodium hyaluronate eyedrops in the treatment of dry eye. Graefes Arch Clin Exp Ophthalmol. 2006;244(1):109–12. 10.1007/s00417-005-0028-1. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
