

Structures of aberrant spliceosome intermediates on their way to disassembly
- PMID: 39833470
- PMCID: PMC12086092
- DOI: 10.1038/s41594-024-01480-7
Structures of aberrant spliceosome intermediates on their way to disassembly
Abstract
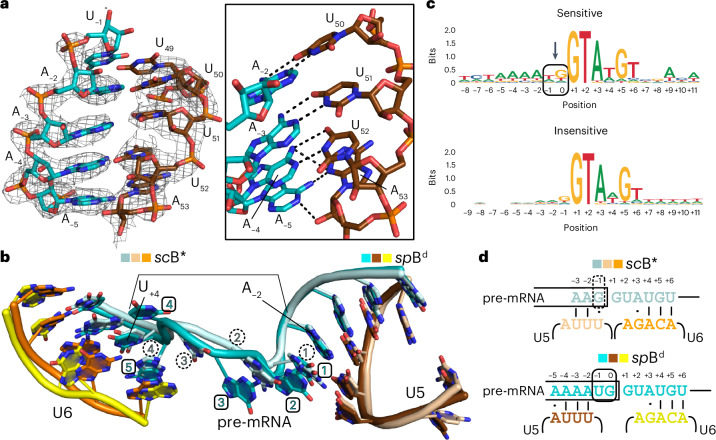
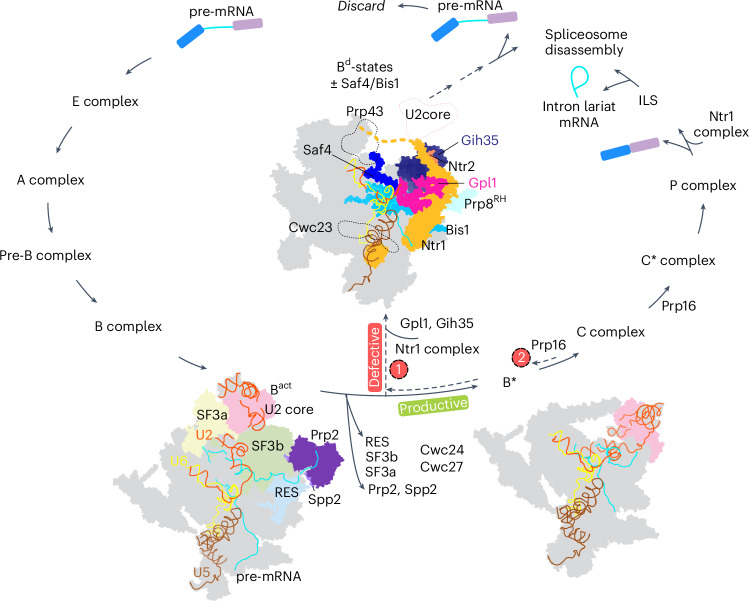
Intron removal during pre-mRNA splicing is of extraordinary complexity and its disruption causes a vast number of genetic diseases in humans. While key steps of the canonical spliceosome cycle have been revealed by combined structure-function analyses, structural information on an aberrant spliceosome committed to premature disassembly is not available. Here, we report two cryo-electron microscopy structures of post-Bact spliceosome intermediates from Schizosaccharomyces pombe primed for disassembly. We identify the DEAH-box helicase-G-patch protein pair (Gih35-Gpl1, homologous to human DHX35-GPATCH1) and show how it maintains catalytic dormancy. In both structures, Gpl1 recognizes a remodeled active site introduced by an overstabilization of the U5 loop I interaction with the 5' exon leading to a single-nucleotide insertion at the 5' splice site. Remodeling is communicated to the spliceosome surface and the Ntr1 complex that mediates disassembly is recruited. Our data pave the way for a targeted analysis of splicing quality control.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures














References
-
- Brody, E. & Abelson, J. The ‘spliceosome’: yeast pre-messenger RNA associates with a 40S complex in a splicing-dependent reaction. Science228, 963–967 (1985). - PubMed
-
- Frendewey, D. & Keller, W. Stepwise assembly of a pre-mRNA splicing complex requires U-snRNPs and specific intron sequences. Cell42, 355–367 (1985). - PubMed
-
- Grabowski, P. J., Seiler, S. R. & Sharp, P. A. A multicomponent complex is involved in the splicing of messenger RNA precursors. Cell42, 345–353 (1985). - PubMed
-
- Shi, Y. Mechanistic insights into precursor messenger RNA splicing by the spliceosome. Nat. Rev. Mol. Cell Biol.18, 655–670 (2017). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
