

Extracellular ATP regulates phagocytic activity, mitochondrial respiration, and cytokine secretion of human astrocytic cells
- PMID: 39833586
- PMCID: PMC12222608
- DOI: 10.1007/s11302-025-10066-x
Extracellular ATP regulates phagocytic activity, mitochondrial respiration, and cytokine secretion of human astrocytic cells
Abstract
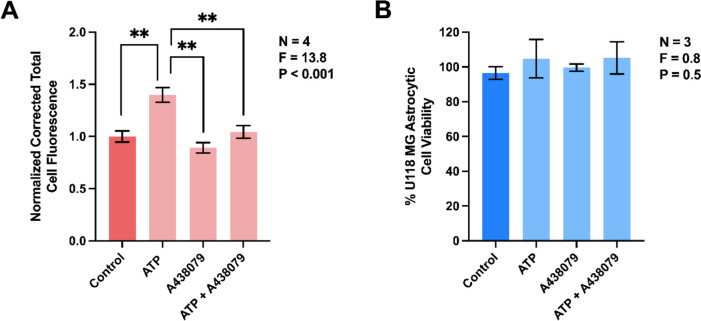
The two main glial cell types of the central nervous system (CNS), astrocytes and microglia, are responsible for neuroimmune homeostasis. Recent evidence indicates astrocytes can participate in removal of pathological structures by becoming phagocytic under conditions of neurodegenerative disease when microglia, the professional phagocytes, are impaired. We hypothesized that adenosine triphosphate (ATP), which acts as damage-associated molecular pattern (DAMP), when released at high concentrations into extracellular space, upregulates phagocytic activity of human astrocytes. This study is the first to measure changes in phagocytic activity and mitochondrial respiration of human astrocytic cells in response to extracellular ATP. We demonstrate that ATP-induced phagocytic activity of U118 MG astrocytic cells is accompanied by upregulated mitochondrial oxidative phosphorylation, which likely supports this energy-dependent process. Application of a selective antagonist A438079 provides evidence identifying astrocytic purinergic P2X7 receptor (P2X7R) as the potential regulator of their phagocytic function. We also report a rapid ATP-induced increase in intracellular calcium ([Ca2+]i), which could serve as regulator of both the phagocytic activity and mitochondrial metabolism, but this hypothesis will need to be tested in future studies. Since ATP upregulates interleukin (IL)-8 secretion by astrocytes but has no effect on their cytotoxicity towards neuronal cells, we conclude that extracellular ATP affects only specific functions of astrocytes. The selectivity of P2X7R-dependent regulation of astrocyte functions by extracellular ATP could allow targeting this receptor-ligand interaction to upregulate their phagocytic function. This could have beneficial outcomes in neurodegenerative disorders, such as Alzheimer's disease, that are characterized by reactive astrocytes and defective phagocytic processes.
Keywords: Astrocytes; Damage-associated molecular pattern (DAMP); Extracellular adenosine triphosphate (ATP); Oxidative phosphorylation; Phagocytosis; Purinergic receptor P2X7.
© 2025. The Author(s), under exclusive licence to Springer Nature B.V.
Conflict of interest statement
Compliance with ethical standards. Ethics approval and consent to participate: All animal experiments were conducted in compliance with the guidelines of the Animal Care Committee at the University of British Columbia, BC, Canada, under protocol A20-0105. Competing interests: The authors declare no competing interests.
Figures




Similar articles
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Mitochondrial damage-associated molecular patterns: Neuroimmunomodulators in central nervous system pathophysiology.Neural Regen Res. 2026 Apr 1;21(4):1322-1338. doi: 10.4103/NRR.NRR-D-24-01459. Epub 2025 Jun 19. Neural Regen Res. 2026. PMID: 40537002 Free PMC article.
-
Management of urinary stones by experts in stone disease (ESD 2025).Arch Ital Urol Androl. 2025 Jun 30;97(2):14085. doi: 10.4081/aiua.2025.14085. Epub 2025 Jun 30. Arch Ital Urol Androl. 2025. PMID: 40583613 Review.
-
MGlu5 Dependent Mitochondrial Translocation of PKCδ: A Mechanism Raising Astrocytic Oxidative Metabolism in Response to Extracellular Glutamate.J Neurochem. 2025 Aug;169(8):e70163. doi: 10.1111/jnc.70163. J Neurochem. 2025. PMID: 40739884 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
