

Emergence of SARS-CoV-2 subgenomic RNAs that enhance viral fitness and immune evasion
- PMID: 39836705
- PMCID: PMC11774490
- DOI: 10.1371/journal.pbio.3002982
Emergence of SARS-CoV-2 subgenomic RNAs that enhance viral fitness and immune evasion
Abstract
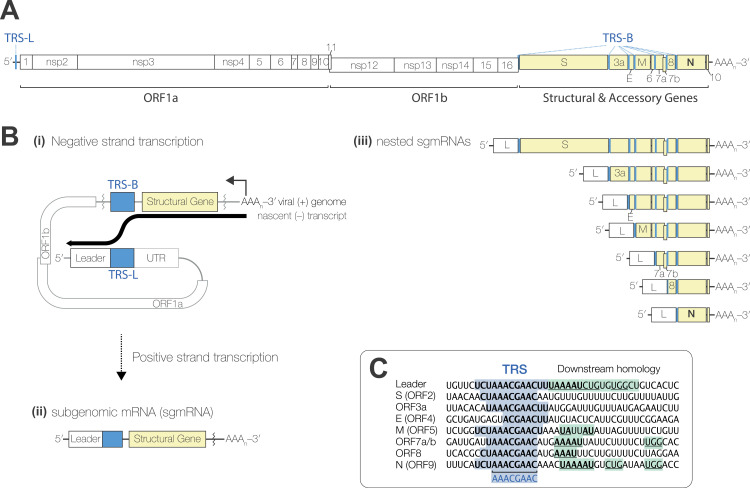
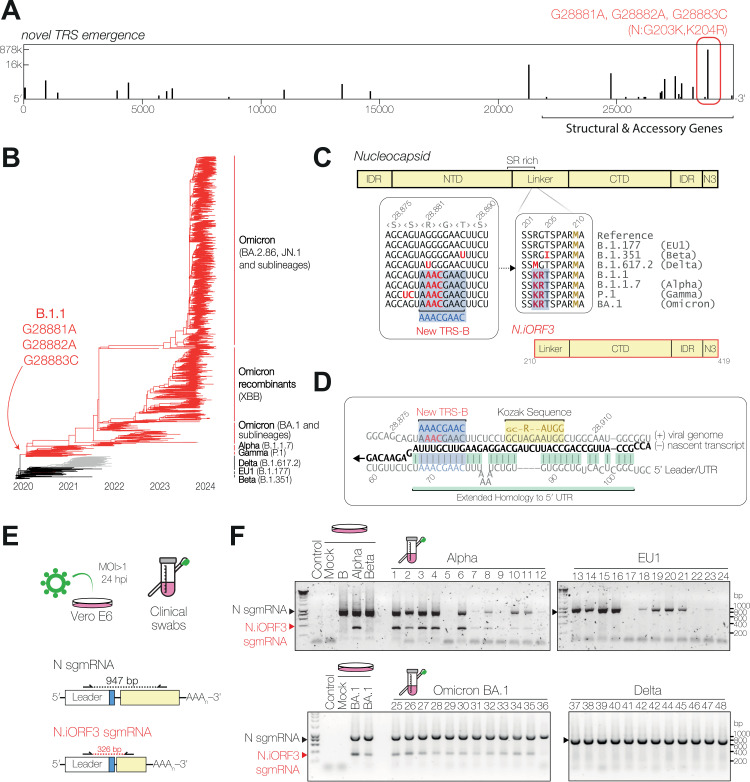
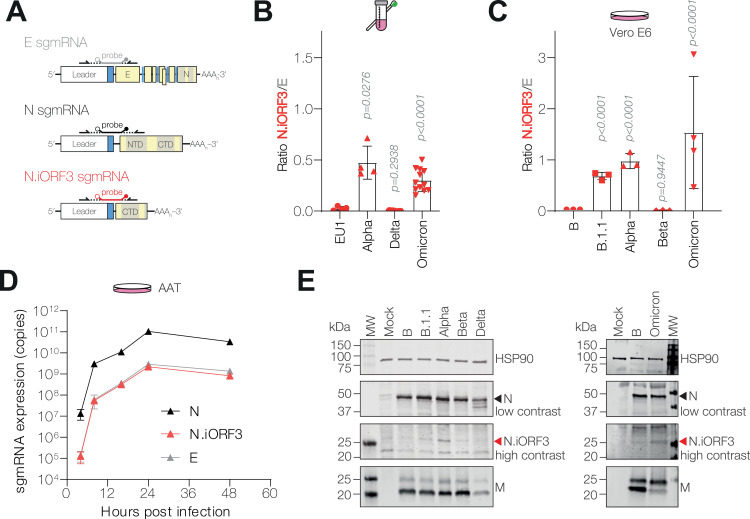
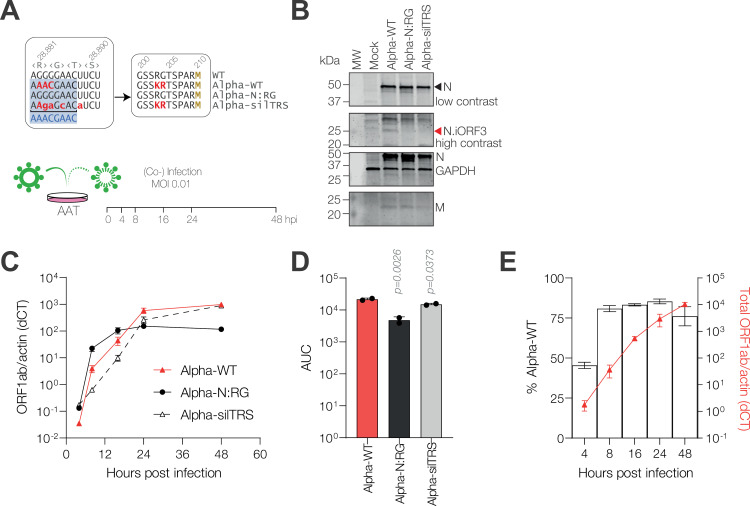
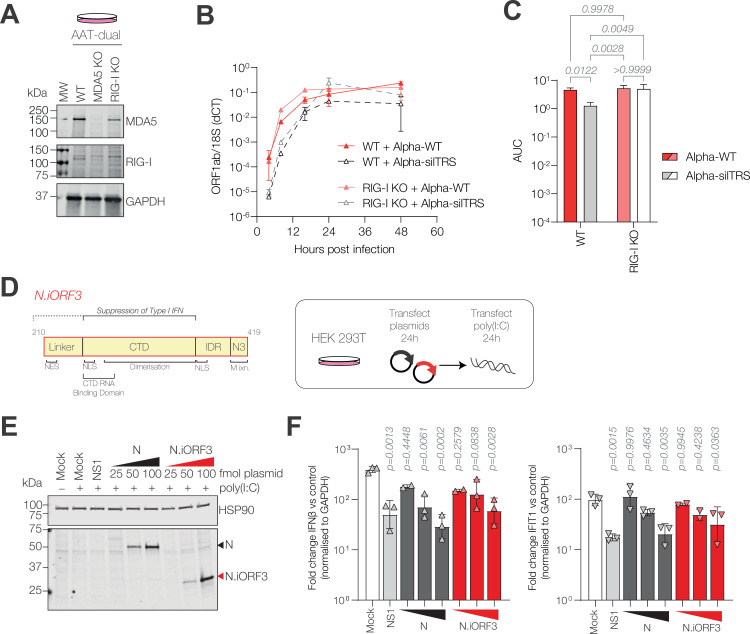
Coronaviruses express their structural and accessory genes via a set of subgenomic RNAs, whose synthesis is directed by transcription regulatory sequences (TRSs) in the 5' genomic leader and upstream of each body open reading frame. In SARS-CoV-2, the TRS has the consensus AAACGAAC; upon searching for emergence of this motif in the global SARS-CoV-2 sequences, we find that it evolves frequently, especially in the 3' end of the genome. We show well-supported examples upstream of the Spike gene-within the nsp16 coding region of ORF1b-which is expressed during human infection, and upstream of the canonical Envelope gene TRS, both of which have evolved convergently in multiple lineages. The most frequent neo-TRS is within the coding region of the Nucleocapsid gene, and is present in virtually all viruses from the B.1.1 lineage, including the variants of concern Alpha, Gamma, Omicron and descendants thereof. Here, we demonstrate that this TRS leads to the expression of a novel subgenomic mRNA encoding a truncated C-terminal portion of Nucleocapsid, which is an antagonist of type I interferon production and contributes to viral fitness during infection. We observe distinct phenotypes when the Nucleocapsid coding sequence is mutated compared to when the TRS alone is ablated. Our findings demonstrate that SARS-CoV-2 is undergoing evolutionary changes at the functional RNA level in addition to the amino acid level.
Copyright: © 2025 Mears et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: While the authors declare no competing interests directly related to this work, CS receives grants from Bristol Myers Squibb, Ono Pharmaceuticals, Boehringer Ingelheim, Roche-Ventana, Pfizer, and Archer Dx; receives personal fees from Genentech, the Sarah Canon Research Institute, Medicxi, Bicycle Therapeutics, GRAIL, Amgen, AstraZeneca, Bristol Myers Squibb, Illumina, GlaxoSmithKline, MSD, and Roche-Ventana; holds stock options in Apogen Biotech, Epic Biosciences, GRAIL, and Achilles Therapeutics; is a member of a scientific advisory board for Bicycle Therapeutics, GRAIL, Relay Therapeutics, SAGA Diagnostics, and Achilles Therapeutics; is a co-founder of Achilles Therapeutics; receives consulting fees from Genentech, Medicxi, MetaboMed, Novartis, the China Innovation Centre of Roche, and the Sarah Cannon Research Institute; and receives honoraria from Amgen, AstraZeneca, Bristol Myers Squibb, Illumina, and Incyte. DLVB receives grants, paid to their institution, from AstraZeneca and GSK related to COVID-19, and is a member of the UK Genotype-to-Phenotype 2 Consortium. All other authors have declared that no competing interests exist.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
